Causal Learning with Neural Assemblies
Pith reviewed 2026-05-07 10:35 UTC · model grok-4.3
The pith
Neural assemblies can learn causal directionality using only their built-in operations of projection, local plasticity control, and sparse winner selection.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
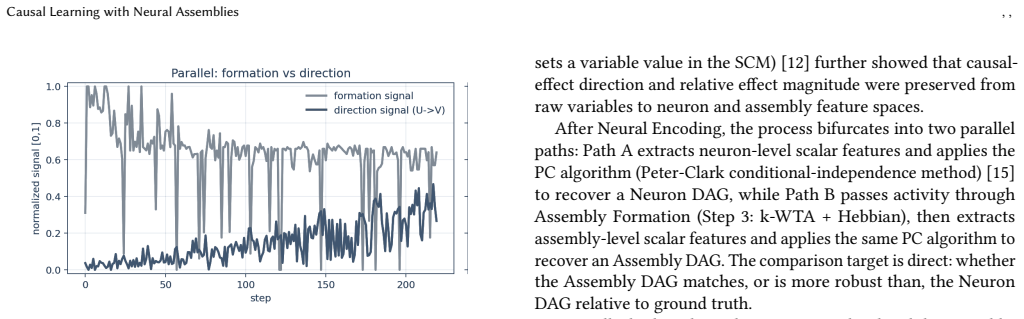
The inherent operations of neural assemblies—projection, local plasticity control, and sparse winner selection—are sufficient for directional learning. DIRECT co-activates source and target assemblies under an adaptive gain schedule to internalize directed relations, yielding perfect structural recovery under supervised known-structure conditions across domains. The resulting causal claims are auditable because they rest on synaptic-strength asymmetry and functional propagation overlap rather than opaque global optimization.
What carries the argument
DIRECT (DIRectional Edge Coupling/Training), a local co-activation rule that pairs source and target assemblies under an adaptive gain schedule to produce measurable synaptic asymmetry and unidirectional propagation.
If this is right
- Causal claims become traceable to specific neural winners and measurable synaptic asymmetries rather than global loss surfaces.
- The same local operations that support classification and planning can now support directional causal graphs.
- Perfect recovery of known causal structures is achieved across multiple domains without backpropagation.
- The framework supplies an explicit bridge between spike-based local dynamics and formal causal semantics.
Where Pith is reading between the lines
- The same local rule could be tested in neuromorphic hardware where global backpropagation is unavailable.
- If the adaptive gain schedule can be made fully unsupervised, the method might extend to causal discovery from observational data alone.
- Disrupting sparse winner selection while preserving co-activation should selectively abolish directional learning while leaving undirected associations intact.
Load-bearing premise
Co-activating source and target assemblies under an adaptive gain schedule is enough to embed directed relations that pass both synaptic-asymmetry and functional-propagation checks.
What would settle it
Run the co-activation schedule on a known directed edge and observe whether the forward synaptic weights remain statistically indistinguishable from the reverse weights or whether activity from the target assembly fails to propagate back to the source more reliably than the reverse.
Figures









read the original abstract
Can Neural Assemblies -- groups of neurons that fire together and strengthen through co-activation -- learn the direction of causal influence between variables? While established as a computationally general substrate for classification, parsing, and planning, neural assemblies have not yet been shown to internalize causal directionality. We demonstrate that the inherent operations of neural assemblies -- projection, local plasticity control, and sparse winner selection -- are sufficient for directional learning. We introduce DIRECT (DIRectional Edge Coupling/Training), a mechanism that co-activates source and target assemblies under an adaptive gain schedule to internalize directed relations. Unlike backpropagation-based methods, DIRECT relies solely on local plasticity, making the resulting causal claims auditable at the mechanism level. Our findings are verified through a dual-readout validation strategy: (i) synaptic-strength asymmetry, measuring the emergent weight gap between forward and reverse links, and (ii) functional propagation overlap, quantifying the reliability of directional signal flow. Across multiple domains, the framework achieves perfect structural recovery under a supervised, known-structure setting. These results establish neural assemblies as an auditable bridge between biologically plausible dynamics and formal causal models, offering an "explainable by design" framework where causal claims are traceable to specific neural winners and synaptic asymmetries.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces DIRECT, a mechanism enabling neural assemblies to internalize directed causal relations via projection, local plasticity control, and sparse winner selection. Source and target assemblies are co-activated under an adaptive gain schedule derived from known causal structure; the resulting weights are validated by two readouts—emergent synaptic-strength asymmetry and directional propagation overlap—yielding perfect structural recovery in a supervised, known-structure regime. The work positions this as a biologically local, auditable alternative to backpropagation for causal modeling.
Significance. If the central mechanism proves robust beyond the supervised regime, the result would supply a concrete, locally computable bridge between neural-assembly dynamics and formal causal graphs, with built-in auditability at the level of individual synaptic asymmetries and winner selections. The dual-readout validation and emphasis on parameter-light local plasticity constitute genuine strengths that could inform both computational neuroscience and explainable causal AI.
major comments (2)
- [Abstract] Abstract: the claim that neural-assembly operations are 'sufficient for directional learning' of causal influence is load-bearing for the title and opening question, yet the reported experiments operate exclusively in a supervised known-structure setting in which the adaptive gain schedule is set using prior knowledge of which assemblies are sources versus targets. This setup demonstrates encoding of externally supplied directions rather than inference of unknown directions from observational data, directly limiting the scope of the central claim.
- [Validation strategy] Validation strategy (abstract and methods): both synaptic-strength asymmetry and functional propagation overlap are defined with respect to the same co-activation schedule used to train the weights. This creates a moderate risk that the reported directionality is partly tautological with the training rule rather than an independent test of learned causality; explicit controls or an observational-data ablation are needed to substantiate the dual-readout as evidence of genuine directional internalization.
minor comments (2)
- The abstract states 'across multiple domains' without enumeration; the results section should list the domains, any domain-specific parameter choices, and the precise criteria for 'perfect structural recovery' (e.g., exact recovery rate, error bars, exclusion rules).
- The adaptive gain schedule is listed as the sole free parameter; its functional form, initialization, and adaptation rule should be given explicitly (ideally with pseudocode or an equation) to support reproducibility.
Simulated Author's Rebuttal
Thank you for the detailed and constructive feedback on our manuscript. We have carefully considered the major comments and provide point-by-point responses below. We believe these revisions will clarify the scope and strengthen the validation of our proposed DIRECT mechanism.
read point-by-point responses
-
Referee: [Abstract] Abstract: the claim that neural-assembly operations are 'sufficient for directional learning' of causal influence is load-bearing for the title and opening question, yet the reported experiments operate exclusively in a supervised known-structure setting in which the adaptive gain schedule is set using prior knowledge of which assemblies are sources versus targets. This setup demonstrates encoding of externally supplied directions rather than inference of unknown directions from observational data, directly limiting the scope of the central claim.
Authors: We agree that the current experiments demonstrate the ability of neural assemblies to encode directed causal relations when provided with a co-activation schedule derived from known causal structure, rather than inferring directions from purely observational data. The claim of sufficiency refers to the operations (projection, local plasticity, sparse winner selection) being adequate to internalize directionality under this supervised regime, which is a necessary first step before tackling unsupervised inference. This is explicitly stated in the manuscript as 'under a supervised, known-structure setting.' To address this, we will revise the abstract and introduction to more precisely delineate the scope, emphasizing that DIRECT provides a mechanism for directional encoding given causal knowledge, positioning it as a building block for future causal inference in neural systems. We will also add a discussion section on extending to observational settings. revision: yes
-
Referee: [Validation strategy] Validation strategy (abstract and methods): both synaptic-strength asymmetry and functional propagation overlap are defined with respect to the same co-activation schedule used to train the weights. This creates a moderate risk that the reported directionality is partly tautological with the training rule rather than an independent test of learned causality; explicit controls or an observational-data ablation are needed to substantiate the dual-readout as evidence of genuine directional internalization.
Authors: We appreciate this concern regarding potential circularity in the validation. The synaptic-strength asymmetry is measured post-training as the difference in weights between forward and reverse connections, which emerges from the asymmetric co-activation schedule. The functional propagation overlap tests whether activation in the source assembly reliably propagates to the target under the learned weights, which is a dynamic test separate from the training phase. However, to mitigate the risk of tautology, we will include additional controls in the revised manuscript: (1) a baseline with symmetric co-activation to show that asymmetry does not emerge without directional scheduling, and (2) an ablation using observational data without the adaptive gain schedule to demonstrate that directionality requires the causal-informed training. These will be added to the methods and results sections to provide stronger evidence of genuine internalization. revision: yes
Circularity Check
Synaptic asymmetry validation is tautological with the imposed co-activation schedule
specific steps
-
self definitional
[Abstract]
"We introduce DIRECT (DIRectional Edge Coupling/Training), a mechanism that co-activates source and target assemblies under an adaptive gain schedule to internalize directed relations. ... Our findings are verified through a dual-readout validation strategy: (i) synaptic-strength asymmetry, measuring the emergent weight gap between forward and reverse links, and (ii) functional propagation overlap, quantifying the reliability of directional signal flow. Across multiple domains, the framework achieves perfect structural recovery under a supervised, known-structure setting."
The adaptive gain schedule explicitly designates which assemblies are co-activated as source vs. target; local plasticity then strengthens only the forward synapses. The measured 'emergent' synaptic-strength asymmetry is therefore the direct, forced outcome of selective co-activation rather than an independent verification that directionality has been learned; the validation confirms the training rule's effect by construction.
full rationale
The paper's core demonstration is that local Hebbian plasticity under a known-structure co-activation schedule produces asymmetric weights and directional propagation. This is shown to work, but the dual-readout metrics directly measure the consequences of the experimenter-supplied source/target schedule rather than an independent test of direction discovery. The mechanism itself (projection, local plasticity, winner selection) is non-circular and auditable, yet the interpretation of 'directional learning' and 'structural recovery' reduces to successful encoding of externally dictated directions. No self-citation chains or ansatzes are load-bearing; the circularity is limited to the validation step being by construction.
Axiom & Free-Parameter Ledger
free parameters (1)
- adaptive gain schedule
axioms (1)
- domain assumption Projection, local plasticity control, and sparse winner selection are sufficient to internalize directed relations when paired with the DIRECT schedule.
invented entities (1)
-
DIRECT mechanism
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Guo-Qiang Bi and Mu-Ming Poo. 1998. Synaptic modifications in cultured hippocampal neurons: dependence on spike timing, synaptic strength, and postsynaptic cell type.Journal of Neuroscience18, 24 (1998), 10464–10472. doi:10.1523/JNEUROSCI.18-24-10464.1998
-
[2]
György Buzsáki. 2010. Neural syntax: cell assemblies, synapsembles, and readers. Neuron68, 3 (2010), 362–385. doi:10.1016/j.neuron.2010.10.023
-
[3]
Max Dabagia, Daniel Mitropolsky, Christos Papadimitriou, and Santosh Vempala
-
[4]
arXiv:2406.07715 [q-bio.NC] doi:10.48550/arXiv.2406.07715
Coin-flipping in the brain: statistical learning with neuronal assemblies. arXiv:2406.07715 [q-bio.NC] doi:10.48550/arXiv.2406.07715
-
[5]
Max Dabagia, Christos Papadimitriou, and Santosh Vempala. 2022. Neural As- semblies of neurons learn to classify well-separated distributions. InProceedings of The 35th Conference on Learning Theory (Proceedings of Machine Learning Research, Vol. 178). PMLR, Online, 4705–4744. https://proceedings.mlr.press/v1 78/dabagia22a.html
2022
-
[6]
Max Dabagia, Christos Papadimitriou, and Santosh Vempala. 2025. Computation with sequences of assemblies in a model of the brain.Neural Computation37, 1 (2025), 193–233. doi:10.1162/neco_a_01720
-
[7]
Francesco d’Amore, Daniel Mitropolsky, Pierluigi Crescenzi, Emanuele Natale, and Christos Papadimitriou. 2022. Planning with biological neurons and synapses. https://aaai.org/papers/00021- planning- with- biological- neurons- and- synapses/ In Proceedings of the AAAI Conference on Artificial Intelligence
2022
- [8]
-
[9]
Jakub Kuzilek, Martin Hlosta, and Zdenek Zdrahal. 2017. Open University Learn- ing Analytics dataset.Scientific Data4 (2017), 170171. doi:10.1038/sdata.2017.171
-
[10]
Henry Markram, Joachim Lübke, Michael Frotscher, and Bert Sakmann. 1997. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science275, 5297 (1997), 213–215. doi:10.1126/science.275.5297.213
-
[11]
Daniel Mitropolsky, Michael Collins, and Christos H. Papadimitriou. 2021. A biologically plausible parser.Transactions of the Association for Computational Linguistics9 (2021), 1374–1388. doi:10.1162/tacl_a_00432
-
[12]
Brain com- putation by assemblies of neurons,
Christos H. Papadimitriou, Santosh Vempala, Daniel Mitropolsky, Michael Collins, and Wolfgang Maass. 2020. Brain computation by neural assemblies of neurons. Proceedings of the National Academy of Sciences117, 25 (2020), 14464–14472. doi:10.1073/pnas.2001893117
-
[13]
Cambridge University Press, 2nd edi- tion, 2009
Judea Pearl. 2009.Causality: Models, Reasoning, and Inference(2nd ed.). Cambridge University Press, Cambridge, UK. doi:10.1017/CBO9780511803161
-
[14]
2017.Elements of Causal Inference: Foundations and Learning Algorithms
Jonas Peters, Dominik Janzing, and Bernhard Schölkopf. 2017.Elements of Causal Inference: Foundations and Learning Algorithms. MIT Press, Cambridge, MA, USA. https://direct.mit.edu/books/book/4376/Elements-of-Causal-Inference
2017
-
[15]
Sen Song, Kenneth D. Miller, and L. F. Abbott. 2000. Competitive Hebbian learning through spike-timing-dependent synaptic plasticity.Nature Neuroscience3, 9 (2000), 919–926. doi:10.1038/78829
-
[16]
2000.Causation, Prediction, and Search(2nd ed.)
Peter Spirtes, Clark Glymour, and Richard Scheines. 2000.Causation, Prediction, and Search(2nd ed.). MIT Press, Cambridge, MA, USA. https://direct.mit.edu/b ooks/book/2605/Causation-Prediction-and-Search
2000
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.