Recognition: 4 theorem links
· Lean TheoremModeling sequential cognitive states via population level cortical dynamics
Pith reviewed 2026-05-08 18:29 UTC · model grok-4.3
The pith
Neural networks approximating dynamics with heteroclinic cycles produce periodic trajectories that closely follow those cycles in an interpretable high-dimensional neural-field model.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
We first show that spatial-discrete neural-field equations with biologically realistic equilibria cannot support heteroclinic cycles. Heteroclinic dynamics often arise in Lotka-Volterra-type systems, but these do not directly correspond to neuronal processes. We use a version of the Universal Approximation Theorem to approximate any target dynamics by a neural network interpretable as a high-dimensional Amari-type neural-field system. When the target dynamics contains a heteroclinic cycle, the approximating vector field generates a periodic trajectory that closely follows the heteroclinic connection. As a case study, we consider the cognitive processes underlying focused-attention meditation
What carries the argument
The neural-network approximant obtained via the Universal Approximation Theorem, interpreted as a high-dimensional Amari-type neural-field system whose equilibria and connections retain population-level meaning.
If this is right
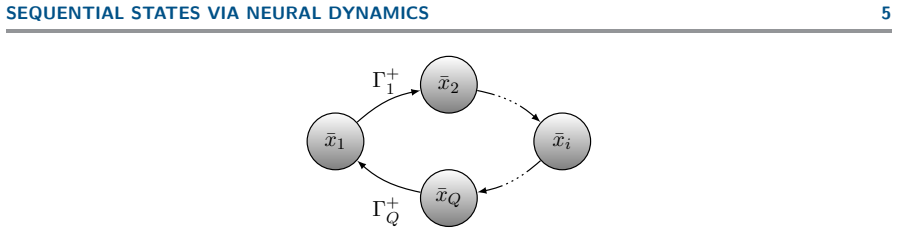
- Sequential transitions among cognitive states arise as periodic trajectories in the approximating neural-field dynamics.
- The model reproduces the cognitive processes underlying focused-attention meditation through population-level cortical dynamics.
- Any target dynamics containing a heteroclinic cycle admits an approximating vector field that tracks the cycle periodically.
- Equilibria and connections in the neural-field system retain biological interpretability at the population level.
Where Pith is reading between the lines
- The technique could extend to modeling other sequential cognitive processes such as decision sequences or memory chains.
- Empirical checks against brain imaging data during meditation tasks would test how well the periodic trajectories match observed activity.
- High-dimensional neural fields may embed low-dimensional heteroclinic structures to support a wide range of sequential brain patterns.
Load-bearing premise
The neural-network approximant obtained via the Universal Approximation Theorem can be interpreted as a biologically plausible high-dimensional Amari-type neural-field system whose equilibria and connections retain meaning at the population level.
What would settle it
A concrete numerical simulation of the approximating neural network for a known target system containing a heteroclinic cycle, verifying whether it generates a periodic trajectory that closely follows the connection.
Figures






read the original abstract
In this work, we present a mathematical model for cyclic and sequential patterns of brain activity, combining heteroclinic dynamics with discrete neural-field models. We first show that spatial-discrete neural-field equations with biologically realistic equilibria cannot support heteroclinic cycles. On the other hand, heterocline dynamics often arise in Lotka-Volterra-type systems, but these equations do not directly correspond to neuronal processes. To address this, we use a version of the Universal Approximation Theorem to approximate any target dynamics by a neural network interpretable as a high-dimensional Amari-type neural-field system. When the target dynamics contains a heteroclinic cycle, the approximating vector field generates a periodic trajectory that closely follows the heteroclinic connection. As a case study, we consider the cognitive processes underlying focused-attention meditation. We show how the model reproduces sequential transitions among cognitive states and we conclude providing a neural interpretation of the approximating dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that spatially discrete neural-field equations with biologically realistic equilibria cannot support heteroclinic cycles, while Lotka-Volterra systems can but lack direct neuronal interpretation. It invokes a version of the Universal Approximation Theorem to show that any target vector field (including those with heteroclinic cycles) can be approximated by a high-dimensional Amari-type neural-field system realized as a neural network. When the target contains a heteroclinic cycle, the approximating system is asserted to generate a periodic trajectory that closely follows the heteroclinic connections. This framework is applied as a case study to model sequential transitions among cognitive states during focused-attention meditation, with a neural interpretation of the resulting dynamics.
Significance. If the central approximation result can be made rigorous with explicit error control, the work would provide a useful bridge between abstract dynamical-systems models of sequential cognition and population-level cortical dynamics interpretable as neural fields. The negative result on discrete neural fields and the explicit neural-network realization are constructive elements that could aid model-building in cognitive neuroscience, though the absence of quantitative bounds on long-time behavior limits immediate applicability.
major comments (2)
- [Abstract / main approximation theorem] Abstract and the statement of the main approximation result: the claim that the approximating vector field 'generates a periodic trajectory that closely follows the heteroclinic connection' is not supported by quantitative error bounds, stability analysis under perturbation, or invocation of shadowing/persistence results. UAT guarantees uniform approximation on compact sets, but heteroclinic orbits spend arbitrarily long times near equilibria; residual errors can alter passage times or destroy the connection structure, and no such control is provided.
- [Case study / meditation application] Case-study section on focused-attention meditation: the manuscript asserts that the model reproduces sequential transitions among cognitive states but provides no verification that the periodic orbit of the approximant preserves the original heteroclinic timing or connection structure; without this, the neural interpretation of the approximating dynamics rests on an unverified shadowing property.
minor comments (2)
- [Model formulation] The precise statement of the neural-field equations (Amari-type) and the embedding of the neural-network weights into the population-level parameters could be clarified with an explicit mapping between the UAT approximant and the biological parameters.
- [Interpretation section] A brief discussion of how the equilibria of the approximant relate to the original saddle points would improve readability and strengthen the biological plausibility argument.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive feedback on our manuscript. The concerns about the lack of quantitative control on the approximation for heteroclinic dynamics and the verification in the case study are well-taken. We will revise the manuscript to temper the claims, add explicit caveats, and clarify the scope of the results. Point-by-point responses follow.
read point-by-point responses
-
Referee: [Abstract / main approximation theorem] Abstract and the statement of the main approximation result: the claim that the approximating vector field 'generates a periodic trajectory that closely follows the heteroclinic connection' is not supported by quantitative error bounds, stability analysis under perturbation, or invocation of shadowing/persistence results. UAT guarantees uniform approximation on compact sets, but heteroclinic orbits spend arbitrarily long times near equilibria; residual errors can alter passage times or destroy the connection structure, and no such control is provided.
Authors: We agree that the Universal Approximation Theorem provides only uniform approximation on compact sets and does not by itself control long-time behavior along heteroclinic orbits, where trajectories linger near equilibria. The original phrasing in the abstract and theorem statement overstated the closeness of the resulting periodic orbit without supporting analysis. In the revision we will replace the claim with a more precise statement that the approximant reproduces the target vector field to arbitrary accuracy on compact subsets of phase space (away from equilibria) and that numerical evidence suggests qualitatively similar sequential transitions; we will add a dedicated paragraph discussing the absence of shadowing or persistence guarantees and the potential for residual errors to affect passage times. This change will be incorporated in the next version. revision: yes
-
Referee: [Case study / meditation application] Case-study section on focused-attention meditation: the manuscript asserts that the model reproduces sequential transitions among cognitive states but provides no verification that the periodic orbit of the approximant preserves the original heteroclinic timing or connection structure; without this, the neural interpretation of the approximating dynamics rests on an unverified shadowing property.
Authors: The case study is intended as an illustrative example rather than a quantitatively validated reproduction. We acknowledge that no explicit check of timing preservation or connection structure is supplied, so the neural interpretation rests on the vector-field approximation alone. In the revised manuscript we will insert a clarifying paragraph stating that the reproduction is qualitative, note the lack of rigorous shadowing results as a limitation, and indicate that future numerical studies could compare passage times between the target and approximant. This addresses the referee's point directly. revision: yes
Circularity Check
No significant circularity; derivation relies on external UAT without self-referential reduction
full rationale
The paper's core step invokes the standard Universal Approximation Theorem to construct a neural-network approximant interpretable as a high-dimensional Amari neural field from an arbitrary target vector field (including one with a heteroclinic cycle). This is not a fitted parameter renamed as a prediction, nor a self-definition, nor dependent on a load-bearing self-citation. The subsequent claim that the approximant generates a periodic trajectory shadowing the heteroclinic connection is a separate mathematical assertion about the approximation's global behavior; it does not reduce by construction to the input target dynamics or to any internal fitting. No equations or steps in the provided abstract and description exhibit the enumerated circularity patterns. The derivation remains self-contained against external benchmarks such as the UAT.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math Universal Approximation Theorem for neural networks
Lean theorems connected to this paper
-
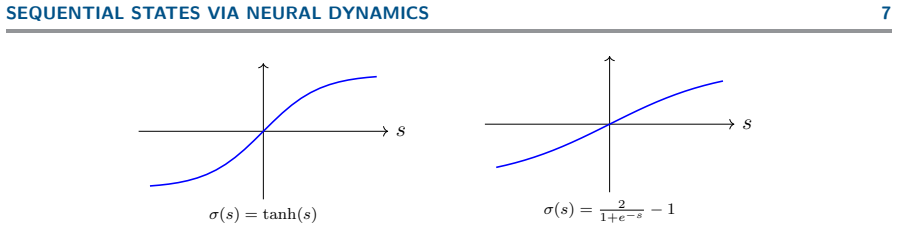
IndisputableMonolith/Cost (Jcost = ½(x+x⁻¹)−1)washburn_uniqueness_aczel unclear˙x = −x + σ(Wx + b) ... σ ∈ C¹(R,R), σ(0)=0, σ′(0)=1, σ(R)⊂(−1,1)
-
IndisputableMonolith/Foundation/AlexanderDualityalexander_duality_circle_linking unclearThree-dimensional state space (CN, DMN, SN) chosen by neuroscience of FAM, not derived
-
Foundation period/8-tick modules(no parameter-free period claim) unclearPeriod of the resulting limit cycle estimated as a sum of residence times ti ∼ 1/λ_u^i set by chosen Lotka-Volterra eigenvalues
-
IndisputableMonolith/Foundation/LogicAsFunctionalEquationderivedCost / Translation Theorem unclearUniversal Approximation Theorem used to approximate an arbitrary smooth vector field by −x + Pσ(Wx+b)
Reference graph
Works this paper leans on
-
[1]
V. S. Afraimovich, M. I. Rabinovich, and P. Varona , Heteroclinic contours in neural ensembles and the winnerless competition principle , International Journal of Bifurcation and Chaos, 14 (2004), pp. 1195–1208
2004
-
[2]
V. S. Afraimovich, V. Zhigulin, and M. I. Rabinovich, On the origin of reproducible sequential activ- ity in neural circuits , Chaos: An Interdisciplinary Journal of Nonlinear Science, 14 (2004), pp. 1123– 1129
2004
-
[3]
T. H. Alderson, A. L. Bokde, J. S. Kelso, L. Maguire, and D. Coyle, Metastable neural dynamics underlies cognitive performance across multiple behavioural paradigms , Human brain mapping, 41 (2020), pp. 3212–3234
2020
-
[4]
S. I. Amari , Characteristics of random nets of analog neuron-like elements , IEEE Transactions on Sys- tems, Man, and Cybernetics, 2 (1972), pp. 643–657, https://doi.org/10.1109/TSMC.1972.4309193
-
[5]
Ashwin and C
P. Ashwin and C. Postlethwaite , On designing heteroclinic networks from graphs , Physica D: Non- linear Phenomena, 265 (2013), pp. 26–39
2013
-
[6]
M. V. Bolelli, G. Citti, A. Sarti, and S. W. Zucker , Individuation of 3d perceptual units from neurogeometry of binocular cells, SIAM Journal on Imaging Sciences, 18 (2025), pp. 2605–2640
2025
-
[7]
M. V. Bolelli and D. Prandi , Neural field equations with time-periodic external inputs and some applications to visual processing, Journal of Mathematical Imaging and Vision, 67 (2025), p. 47
2025
-
[8]
P. C. Bressloff, J. D. Cowan, M. Golubitsky, P. J. Thomas, and M. C. Wiener, Geometric visual hallucinations, Euclidean symmetry and the functional architecture of striate cortex , Philosophical Transactions of the Royal Society B: Biological Sciences, 356 (2001), pp. 299–330, https://doi.org/ 10.1098/rstb.2000.0769
-
[9]
Capouskova, M
K. Capouskova, M. L. Kringelbach, and G. Deco , Modes of cognition: Evidence from metastable brain dynamics, Neuroimage, 260 (2022), p. 119489
2022
-
[10]
S. B. Castro and A. M. Rucklidge , Robust heteroclinic cycles in pluridimensions: S. castro et al. , Journal of Nonlinear Science, 35 (2025), p. 80. 28 M. V. BOLELLI, L. GRECO, D. PRANDI
2025
-
[11]
Coombes, P
S. Coombes, P. Beim Graben, R. Potthast, and J. Wright , eds., Neural Fields: Theory and Applications , Springer Berlin Heidelberg, Berlin, Heidelberg, 2014, https://doi.org/10.1007/ 978-3-642-54593-1
2014
-
[12]
J. P. Cunningham and B. M. Yu , Dimensionality reduction for large-scale neural recordings , Nature Neuroscience, 17 (2014), pp. 1500–1509, https://doi.org/10.1038/nn.3776
-
[13]
Cybenko, Approximation by superpositions of a sigmoidal function , Mathematics of control, signals and systems, 2 (1989), pp
G. Cybenko, Approximation by superpositions of a sigmoidal function , Mathematics of control, signals and systems, 2 (1989), pp. 303–314
1989
-
[14]
G. B. Ermentrout and J. D. Cowan, A mathematical theory of visual hallucination patterns. , Biolog- ical cybernetics, 34 (1979), pp. 137–150, https://doi.org/10.1007/BF00336965
-
[15]
Ganesan, E
S. Ganesan, E. Beyer, B. Moffat, N. T. Van Dam, V. Lorenzetti, and A. Zalesky, Focused atten- tion meditation in healthy adults: A systematic review and meta-analysis of cross-sectional functional mri studies, Neuroscience & Biobehavioral Reviews, 141 (2022), p. 104846
2022
-
[16]
W. Hasenkamp, Using first-person reports during meditation to investigate basic cognitive experience , in Meditation–Neuroscientific Approaches and Philosophical Implications, Springer, 2013, pp. 75–93
2013
-
[17]
Hasenkamp and L
W. Hasenkamp and L. W. Barsalou , Effects of meditation experience on functional connectivity of distributed brain networks, Frontiers in human neuroscience, 6 (2012), p. 38
2012
-
[18]
Hasenkamp, C
W. Hasenkamp, C. D. Wilson-Mendenhall, E. Duncan, and L. W. Barsalou , Mind wandering and attention during focused meditation: a fine-grained temporal analysis of fluctuating cognitive states, Neuroimage, 59 (2012), pp. 750–760
2012
-
[19]
A. D. Horchler, K. A. Daltorio, H. J. Chiel, and R. D. Quinn , Designing responsive pattern generators: stable heteroclinic channel cycles for modeling and control , Bioinspiration & biomimetics, 10 (2015), p. 026001
2015
-
[20]
Hornik, M
K. Hornik, M. Stinchcombe, and H. White , Universal approximation of an unknown mapping and its derivatives using multilayer feedforward networks , Neural networks, 3 (1990), pp. 551–560
1990
-
[21]
Huerta and M
R. Huerta and M. Rabinovich, Reproducible sequence generation in random neural ensembles, Physical review letters, 93 (2004), p. 238104
2004
-
[22]
Jeong and C
V. Jeong and C. Postlethwaite, Effect of noise on residence times of a heteroclinic cycle , Dynamical Systems, 38 (2023), pp. 79–101
2023
-
[23]
Krupa, Robust heteroclinic cycles, Journal of Nonlinear Science, 7 (1997), pp
M. Krupa, Robust heteroclinic cycles, Journal of Nonlinear Science, 7 (1997), pp. 129–176
1997
-
[24]
Krupa and I
M. Krupa and I. Melbourne , Asymptotic stability of heteroclinic cycles in systems with symmetry , Ergodic Theory and Dynamical Systems, 15 (1995), pp. 121–147
1995
-
[25]
Laurent, M
G. Laurent, M. Stopfer, R. W. Friedrich, M. I. Rabinovich, A. Volkovskii, and H. D. Abar- banel, Odor encoding as an active, dynamical process: experiments, computation, and theory, Annual review of neuroscience, 24 (2001), pp. 263–297
2001
-
[26]
J. M. Lee and J. M. Lee , Smooth manifolds, Springer, 2003
2003
-
[27]
A. Maier and D. A. Leopold , Binocular rivalry , in Encyclopedia of Neuroscience, M. D. Binder, N. Hirokawa, and U. Windhorst, eds., Springer Berlin Heidelberg, Berlin, Heidelberg, 2009, pp. 391– 394, https://doi.org/10.1007/978-3-540-29678-2 631
-
[28]
K. Nechyporenko, P. Ashwin, and K. Tsaneva-Atanasova, Switching states: Heteroclinic cycles as organising centres of neuronal dynamics , arXiv preprint arXiv:2507.15519, (2025)
-
[29]
Nicolas, B
J. Nicolas, B. R. King, D. L ´evesque, L. Lazzouni, G. Leroux, D. Wang, N. Grossman, S. P. Swinnen, J. Doyon, J. Carrier, et al. , Unraveling the neurophysiological correlates of phase- specific enhancement of motor memory consolidation via slow-wave closed-loop targeted memory re- activation, Nature Communications, 16 (2025), p. 2644
2025
-
[30]
arXiv preprint arXiv:2407.18384 , year=
P. Petersen and J. Zech , Mathematical theory of deep learning , arXiv preprint arXiv:2407.18384, (2024)
-
[31]
Pinkus, Approximation theory of the mlp model in neural networks , Acta numerica, 8 (1999), pp
A. Pinkus, Approximation theory of the mlp model in neural networks , Acta numerica, 8 (1999), pp. 143– 195
1999
-
[32]
Rabinovich, A
M. Rabinovich, A. Volkovskii, P. Lecanda, R. Huerta, H. D. Abarbanel, and G. Laurent , Dynamical encoding by networks of competing neuron groups: winnerless competition, Physical review letters, 87 (2001), p. 068102
2001
-
[33]
M. I. Rabinovich, V. S. Afraimovich, C. Bick, and P. Varona , Information flow dynamics in the brain, Physics of Life Reviews, 9 (2012), pp. 51–73, https://doi.org/10.1016/j.plrev.2011.11.002
-
[34]
M. I. Rabinovich, R. Huerta, P. Varona, and V. S. Afraimovich , Transient cognitive dynamics, SEQUENTIAL STATES VIA NEURAL DYNAMICS 29 metastability, and decision making , PLoS computational biology, 4 (2008), p. e1000072
2008
-
[35]
M. I. Rabinovich, P. Varona, A. I. Selverston, and H. D. Abarbanel , Dynamical principles in neuroscience, Reviews of modern physics, 78 (2006), pp. 1213–1265
2006
-
[36]
Ricard, A
M. Ricard, A. Lutz, and R. J. Davidson , Mind of the meditator , Scientific American, 311 (2014), pp. 38–45
2014
-
[37]
Schwappach, A
C. Schwappach, A. Hutt, and P. Beim Graben , Metastable dynamics in heterogeneous neural fields , Frontiers in systems neuroscience, 9 (2015), p. 97
2015
-
[38]
H. L. Smith , Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Coop- erative Systems, vol. 41 of Mathematical Surveys and Monographs, American Mathematical Society, 1995
1995
-
[39]
Stone and P
E. Stone and P. Holmes , Random perturbations of heteroclinic attractors , SIAM Journal on Applied Mathematics, 50 (1990), pp. 726–743
1990
-
[40]
C. Stringer, M. Pachitariu, N. Steinmetz, M. Carandini, and K. D. Harris , High-dimensional geometry of population responses in visual cortex , Nature, 571 (2019), pp. 361–365, https://doi.org/ 10.1038/s41586-019-1346-5
-
[41]
Tamekue, D
C. Tamekue, D. Prandi, and Y. Chitour , A Mathematical Model of the Visual MacKay Effect , SIAM Journal on Applied Dynamical Systems, 23 (2024), pp. 2138–2178, https://doi.org/10.1137/ 23M1616686
2024
-
[42]
C. Tamekue, D. Prandi, and Y. Chitour , Reproducibility via neural fields of visual illusions induced by localized stimuli , Discrete and Continuous Dynamical Systems - B, 30 (2025), pp. 1441–1471, https://doi.org/10.3934/dcdsb.2024135
-
[43]
M. W. van Es, C. Higgins, C. Gohil, A. J. Quinn, D. Vidaurre, and M. W. Woolrich , Large- scale cortical functional networks are organized in structured cycles , Nature Neuroscience, 28 (2025), pp. 2118–2128
2025
-
[44]
H. R. Wilson and J. D. Cowan, Excitatory and inhibitory interactions in localized populations of model neurons, Biophysical journal, 12 (1972), pp. 1–24
1972
-
[45]
H. R. Wilson and J. D. Cowan, Excitatory and inhibitory interactions in localized populations of model neurons, Biophysical Journal, 12 (1972), pp. 1–24, https://doi.org/10.1016/S0006-3495(72)86068-5
-
[46]
H. R. Wilson and J. D. Cowan , A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue , Kybernetik, 13 (1973), pp. 55–80
1973
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.