Recognition: unknown
Loop Extrusion Reversal by Condensin Motor is Mediated by Catch Bonds
Pith reviewed 2026-05-08 15:49 UTC · model grok-4.3
The pith
Condensin reverses DNA loop extrusion through a force-sensitive intermediate state that acts like catch bonds.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
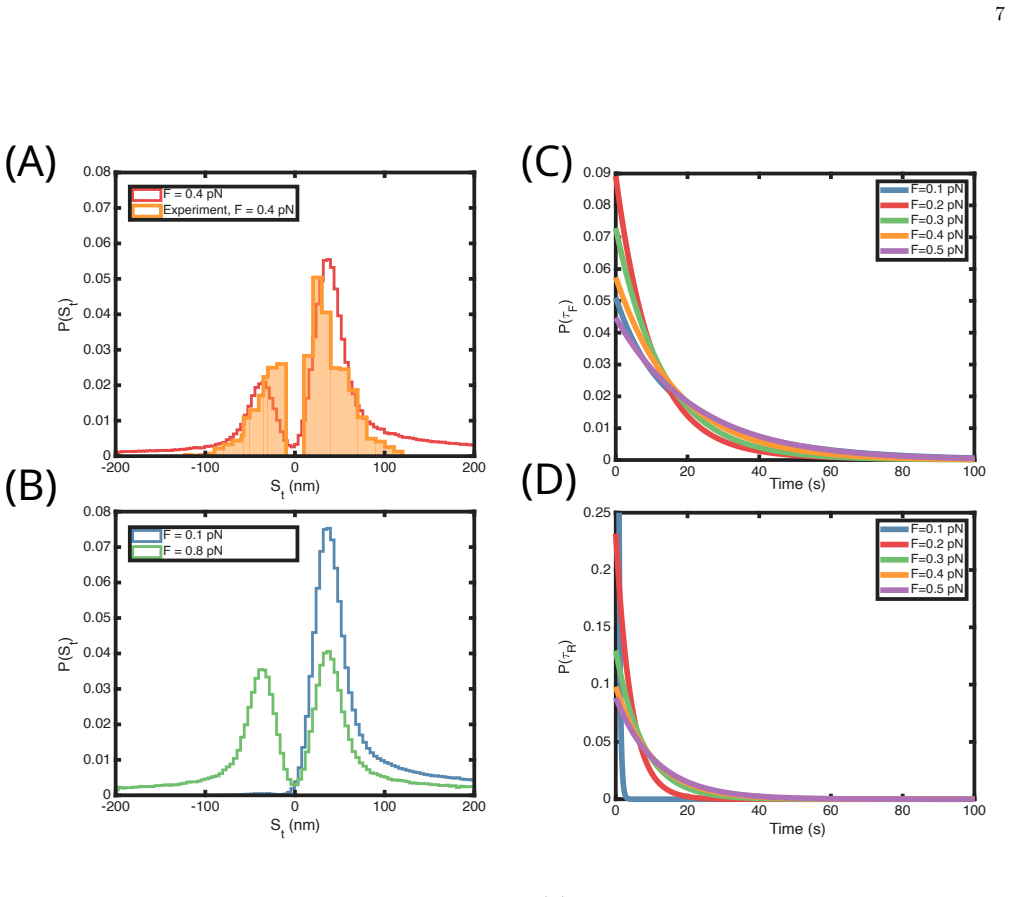
Using a stochastic kinetic model based on the scrunching mechanism, the calculations quantitatively account for the measured force-dependent step size and dwell time distributions in both the forward and backward directions. By postulating the existence of an intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step, the model predicts that its lifetime increases as the external mechanical force increases till a critical value and subsequently decreases at higher forces. The surprising finding of lifetime increase in an active motor at sub-piconewton forces is the characteristic of catch bonds.
What carries the argument
The intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step, whose lifetime exhibits non-monotonic dependence on external force.
If this is right
- The model quantitatively reproduces the force-dependent step sizes and dwell times observed for both forward and reverse extrusion steps.
- The lifetime of the intermediate state is predicted to increase with force until a critical value and then decrease.
- Catch-bond behavior appears in an active motor, expanding the known ways mechanical force can regulate loop extrusion.
- Mechanical forces are highlighted as regulators of genome organization through condensin activity.
Where Pith is reading between the lines
- The same catch-bond intermediate may exist in other SMC complexes that organize chromosomes.
- Local cellular forces could dynamically switch extrusion direction without changing the motor itself.
- Structural or single-molecule studies could test for the predicted intermediate state directly.
- The mechanism offers a way for mechanical stress to control genome folding patterns in living cells.
Load-bearing premise
The existence of an intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step, introduced to explain both directions without independent structural or kinetic evidence.
What would settle it
Direct measurement of the lifetime of the proposed intermediate state as a function of applied force, which should increase up to a critical value and then decrease at higher forces.
Figures




read the original abstract
Structural Maintenance Complexes (SMC) are energy consuming motors that are important in folding the genome by loop extrusion (LE) in all stages of the cell cycle. Single molecule magnetic tweezer pulling experiments have revealed that condensin, a member of the SMC family involved in mitosis, takes occasional backward steps, thus coughing up the gains in the length of the extruded loop. To reveal the mechanism of the forward and backward steps simultaneously, we developed a theory using the stochastic kinetic model and the scrunching mechanism for LE. The calculations quantitatively account for the measured force-dependent step size and dwell time distributions in both the directions. By postulating the existence of an intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step, we predict that its lifetime increases as the external mechanical force increases till a critical value and subsequently decreases at higher forces. The surprising finding of lifetime increase in an active motor, at sub-piconewton forces, is the characteristic of catch bonds, known in force-induced rupture of several passive protein complexes. The identification of catch bond-like states in condensin not only expands our understanding of LE but also highlights the significance of mechanical forces in regulating genome organization.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops a stochastic kinetic model of condensin loop extrusion based on a scrunching mechanism. By introducing an intermediate state in the ATP-driven cycle that is poised to commit to either a forward or backward step, the model is claimed to quantitatively reproduce experimental force-dependent step sizes and dwell-time distributions in both directions. The same state is used to predict that the lifetime of this intermediate increases with applied force up to a critical value and then decreases, a non-monotonic dependence interpreted as catch-bond behavior in an active motor.
Significance. If the quantitative fits can be reproduced with full model transparency and the postulated intermediate state receives independent structural or kinetic support, the work would link mechanical force to bidirectional loop extrusion and suggest that catch-bond-like regulation operates in SMC motors, with implications for genome folding. The absence of explicit equations, parameters, and orthogonal validation currently prevents assessment of whether these results are robust or circular.
major comments (3)
- [Abstract and model section] Abstract and model section: the claim that the calculations 'quantitatively account for the measured force-dependent step size and dwell time distributions in both the directions' cannot be evaluated because the manuscript supplies neither the explicit kinetic rate equations, the numerical values of the free parameters (kinetic rate constants), nor the fitting procedure used to match the data.
- [Results on intermediate-state lifetime] Results on intermediate-state lifetime: the predicted non-monotonic force dependence of the intermediate-state lifetime is generated by the same postulated state that was introduced to explain backward steps, rendering the result circular with respect to the modeling assumptions rather than an independent test. No structural (cryo-EM) or orthogonal kinetic (FRET) evidence is cited for the existence or force sensitivity of this state.
- [Discussion] Discussion: the existence of an 'intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step' is the load-bearing axiom for both the bidirectional-motion account and the catch-bond prediction; without independent justification, the central claims rest on an ad-hoc entity whose removal would eliminate the reported force dependence.
minor comments (1)
- A diagram explicitly labeling the kinetic states, transitions, and force-dependent rates would improve clarity of the stochastic model.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments on our manuscript. We address each major comment point by point below, providing clarifications on the model and indicating where revisions have been made to improve transparency and justification.
read point-by-point responses
-
Referee: [Abstract and model section] Abstract and model section: the claim that the calculations 'quantitatively account for the measured force-dependent step size and dwell time distributions in both the directions' cannot be evaluated because the manuscript supplies neither the explicit kinetic rate equations, the numerical values of the free parameters (kinetic rate constants), nor the fitting procedure used to match the data.
Authors: We agree that the main text did not provide sufficient detail on the kinetic equations and parameters. In the revised manuscript, we have added a new subsection in the Methods that presents the full set of stochastic kinetic rate equations for the scrunching-based model, including the explicit forms for forward and backward transitions from the intermediate state. All numerical values of the rate constants are now tabulated, along with a description of the fitting procedure (least-squares minimization against the experimental step-size and dwell-time histograms at multiple forces). These additions enable direct evaluation and reproduction of the quantitative agreement with the bidirectional data. revision: yes
-
Referee: [Results on intermediate-state lifetime] Results on intermediate-state lifetime: the predicted non-monotonic force dependence of the intermediate-state lifetime is generated by the same postulated state that was introduced to explain backward steps, rendering the result circular with respect to the modeling assumptions rather than an independent test. No structural (cryo-EM) or orthogonal kinetic (FRET) evidence is cited for the existence or force sensitivity of this state.
Authors: The intermediate state is introduced to capture the physical requirement, within the scrunching mechanism, that the motor can commit to either direction after ATP hydrolysis. Its parameters are constrained solely by fitting the observed force-dependent step sizes and dwell-time distributions; the non-monotonic lifetime is an emergent prediction from the force dependence of the exit rates from that state and was not used as a fitting target. We have revised the Results and Discussion to explicitly separate the fitting step from the lifetime prediction and to clarify this distinction. We acknowledge the absence of direct cryo-EM or FRET data and have added a paragraph noting this limitation while outlining possible future experiments to test the state's force sensitivity. revision: partial
-
Referee: [Discussion] Discussion: the existence of an 'intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step' is the load-bearing axiom for both the bidirectional-motion account and the catch-bond prediction; without independent justification, the central claims rest on an ad-hoc entity whose removal would eliminate the reported force dependence.
Authors: The intermediate state follows directly from the scrunching mechanism, in which the condensin head domains must form a configuration that can resolve into either forward extrusion or reversal under load. Its inclusion is justified by the model's ability to simultaneously reproduce multiple independent experimental observables (step-size distributions and dwell times in both directions) with a single parameter set. We have expanded the Discussion to provide a more detailed mechanistic derivation of why such a state is required by the scrunching geometry and to discuss how its removal would be inconsistent with the observed reversals. While we agree that additional orthogonal evidence would be desirable, the current data-driven validation supports retaining the state. revision: partial
Circularity Check
Postulated intermediate state generates non-monotonic lifetime prediction by construction
specific steps
-
fitted input called prediction
[Abstract]
"By postulating the existence of an intermediate state in the ATP-driven cycle that is poised to take a forward or a backward step, we predict that its lifetime increases as the external mechanical force increases till a critical value and subsequently decreases at higher forces."
The non-monotonic lifetime dependence is derived from the stochastic kinetic model whose central feature is the postulated intermediate state introduced to fit the observed bidirectional step sizes and dwell-time distributions. The 'prediction' is therefore a direct output of the fitted model rather than an independent derivation from external data or first principles.
full rationale
The paper develops a stochastic kinetic model of the scrunching mechanism that quantitatively accounts for measured force-dependent step sizes and dwell-time distributions. To simultaneously explain forward and backward steps, an intermediate state poised for either direction is postulated. The model then yields a non-monotonic force dependence for the lifetime of this state, presented as a prediction and linked to catch-bond behavior. Because the lifetime is a direct output of the same kinetic scheme and parameters introduced to fit the bidirectional data, the claimed prediction reduces to a consequence of the modeling assumptions rather than an independent result. No orthogonal structural or kinetic evidence for the state is cited, producing partial circularity in the central claim.
Axiom & Free-Parameter Ledger
free parameters (1)
- kinetic rate constants
axioms (2)
- domain assumption Scrunching mechanism underlies loop extrusion
- ad hoc to paper Existence of an intermediate state poised for forward or backward step
invented entities (1)
-
Catch-bond-like intermediate state
no independent evidence
Reference graph
Works this paper leans on
-
[1]
power stroke
to determine their values. The best fit parameters are given in Table II. The fitted and experimental values of the number of forward and reverse steps are shown in Figs. 2(A) and (B) and the dwell time distribution of the forward and reverse steps are shown in Figs. 2(C) and (D). The fits were stable with respect to random starting conditions 4 FIG. 1.Sc...
-
[2]
(rN −r 1)2 + N−1X i=1 (ri+1 −r i)2 # .(13) To preserve translational invariance, we modified the Hamiltonian by tethering the first monomer to the origin giving us, H= 3kBT 2b2
atF= 0.4 pN. for the parameters. The predictions for the dwell time distributions at other values of forces are also plotted (Figs. 3(C) and (D). III. RESULTS Step-size distribution:With the values of the rates in hand, we used the kinetic equations to compute the properties of the loop extrusion model. We first calcu- lated the step-size distributions of...
-
[3]
In the second case (Process 2), aX 1(i+1)→X 0(i+1) step followed by anotherX 0(i+ 1)→X 1(i+ 2) step occurs
is theX 0(i) toX 1(i+ 1) step for which the dwell time distribution is given by, f1(t) =k + 1 e−k+ 1 t (27) The average dwell time in this case is 1/k + 1 . In the second case (Process 2), aX 1(i+1)→X 0(i+1) step followed by anotherX 0(i+ 1)→X 1(i+ 2) step occurs. In this case the dwell time distribution, f2(t)∼ Z t 0 dt′k+ 2 e−(k+ 2 +k− 1 )t′ k+ 1 e−k+ 1...
-
[4]
Uhlmann, SMC complexes: from DNA to chromo- somes, Nature reviews Molecular cell biology17, 399 (2016)
F. Uhlmann, SMC complexes: from DNA to chromo- somes, Nature reviews Molecular cell biology17, 399 (2016)
2016
-
[5]
Nasmyth and C
K. Nasmyth and C. H. Haering, The structure and func- tion of smc and kleisin complexes, Annu. Rev. Biochem. 74, 595 (2005)
2005
-
[6]
Goundaroulis, E
D. Goundaroulis, E. Lieberman Aiden, and A. Stasiak, Chromatin Is Frequently Unknotted at the Megabase Scale, Biophysical Journal118, 2268 (2020)
2020
-
[7]
Hirano, R
T. Hirano, R. Kobayashi, and M. Hirano, Condensins, chromosome condensation protein complexes containing xcap-c, xcap-e and a xenopus homolog of the drosophila barren protein, Cell89, 511–521 (1997)
1997
-
[8]
Michaelis, R
C. Michaelis, R. Ciosk, and K. Nasmyth, Cohesins: Chro- mosomal proteins that prevent premature separation of sister chromatids, Cell91, 35–45 (1997)
1997
-
[9]
Pradhan, T
B. Pradhan, T. Kanno, M. Umeda Igarashi, M. S. Loke, M. D. Baaske, J. S. K. Wong, K. Jeppsson, C. Bj¨ orkegren, and E. Kim, The smc5/6 complex is a dna loop-extruding motor, Nature616, 843 (2023)
2023
-
[10]
Takaki, A
R. Takaki, A. Dey, G. Shi, and D. Thirumalai, Theory and simulations of condensin mediated loop extrusion in dna, Nature Communications12, 5865 (2021)
2021
-
[11]
J. F. Marko, P. De Los Rios, A. Barducci, and S. Gruber, DNA-segment-capture model for loop extrusion by struc- tural maintenance of chromosome (SMC) protein com- plexes, Nucleic acids research47, 6956 (2019)
2019
-
[12]
Hassler, I
M. Hassler, I. A. Shaltiel, and C. H. Haering, Towards a unified model of smc complex function, Current Biology 28, R1266 (2018)
2018
-
[14]
Dekker, C
C. Dekker, C. H. Haering, J.-M. Peters, and B. D. Row- land, How do molecular motors fold the genome?, Science 382, 646 (2023)
2023
-
[15]
Ganji, I
M. Ganji, I. A. Shaltiel, S. Bisht, E. Kim, A. Kalichava, C. H. Haering, and C. Dekker, Real-time imaging of dna loop extrusion by condensin, Science360, 102 (2018)
2018
-
[16]
I. F. Davidson, B. Bauer, D. Goetz, W. Tang, G. Wutz, and J.-M. Peters, DNA loop extrusion by human cohesin, Science366, 1338 (2019)
2019
-
[17]
Y. Kim, Z. Shi, H. Zhang, I. J. Finkelstein, and H. Yu, Human cohesin compacts DNA by loop extrusion, Sci- ence366, 1345 (2019)
2019
-
[19]
Alipour and J
E. Alipour and J. F. Marko, Self-organization of do- main structures by DNA-loop-extruding enzymes, Nu- cleic acids research40, 11202 (2012)
2012
-
[20]
Fudenberg, M
G. Fudenberg, M. Imakaev, C. Lu, A. Goloborodko, N. Abdennur, and L. A. Mirny, Formation of chromo- somal domains by loop extrusion, Cell reports15, 2038 (2016)
2038
-
[21]
Revyakin, C
A. Revyakin, C. Liu, R. H. Ebright, and T. R. Strick, Abortive initiation and productive initiation by rna polymerase involve DNA scrunching, Science314, 1139 (2006)
2006
-
[22]
A. N. Kapanidis, E. Margeat, S. O. Ho, E. Kortkhonjia, S. Weiss, and R. H. Ebright, Initial transcription by RNA polymerase proceeds through a DNA-scrunching mecha- nism, Science314, 1144 (2006)
2006
-
[23]
A. Dey, G. Shi, R. Takaki, and D. Thirumalai, Structural changes in chromosomes driven by multiple condensin motors during mitosis, Cell Reports42(2023)
2023
-
[24]
Golfier, T
S. Golfier, T. Quail, H. Kimura, and J. Brugu´ es, Cohesin and condensin extrude dna loops in a cell cycle-dependent manner, Elife9, e53885 (2020)
2020
-
[25]
R. C. Tolman, The principle of microscopic reversibility, Proceedings of the National Academy of Sciences11, 436 (1925)
1925
-
[26]
M. L. Mugnai, C. Hyeon, M. Hinczewski, and D. Thiru- malai, Theoretical perspectives on biological machines, Reviews of Modern Physics92, 025001 (2020)
2020
-
[27]
A. B. Kolomeisky and M. E. Fisher, Molecular motors: a theorist’s perspective, Annu. Rev. Phys. Chem.58, 675 (2007)
2007
-
[28]
Ryu, S.-H
J.-K. Ryu, S.-H. Rah, R. Janissen, J. W. Kerssemakers, A. Bonato, D. Michieletto, and C. Dekker, Condensin extrudes DNA loops in steps up to hundreds of base pairs that are generated by ATP binding events, Nucleic acids research50, 820 (2022)
2022
-
[29]
T. Yamamoto and H. Schiessel, Osmotic mechanism of the loop extrusion process, Physical Review E96, 10.1103/physreve.96.030402 (2017)
-
[30]
C. Brackley, J. Johnson, D. Michieletto, A. Morozov, M. Nicodemi, P. Cook, and D. Marenduzzo, Nonequilib- rium chromosome looping via molecular slip links, Phys- ical Review Letters119, 10.1103/physrevlett.119.138101 (2017)
-
[31]
C. A. Brackley, J. Johnson, D. Michieletto, A. N. Mo- rozov, M. Nicodemi, P. R. Cook, and D. Marenduzzo, Extrusion without a motor: a new take on the loop ex- trusion model of genome organization, Nucleus9, 95–103 (2018)
2018
-
[32]
Bonato and D
A. Bonato and D. Michieletto, Three-dimensional loop extrusion, Biophysical Journal120, 5544–5552 (2021)
2021
-
[33]
T. L. Higashi, G. Pobegalov, M. Tang, M. I. Molodtsov, and F. Uhlmann, A brownian ratchet model for dna loop extrusion by the cohesin complex, eLife10, 10.7554/elife.67530 (2021)
-
[34]
Dembo, D
M. Dembo, D. Torney, K. Saxman, and D. Hammer, The reaction-limited kinetics of membrane-to-surface ad- hesion and detachment, Proceedings of the Royal Society of London. Series B. Biological Sciences234, 55 (1988)
1988
-
[35]
Barsegov and D
V. Barsegov and D. Thirumalai, Dynamics of unbinding of cell adhesion molecules: Transition from catch to slip bonds, Proc. Natl. Acad. Sci. USA102, 1835 (2005)
2005
-
[36]
Barsegov and D
V. Barsegov and D. Thirumalai, Dynamic competition between catch and slip bonds in selectins bound to lig- ands, The Journal of Physical Chemistry B110, 26403 (2006)
2006
-
[37]
Thomas, V
W. Thomas, V. Vogel, and E. Sokurenko, Biophysics of catch bonds, Annu. Rev. Biophys.37, 399 (2008)
2008
-
[38]
B. T. Marshall, M. Long, J. W. Piper, T. Yago, R. P. McEver, and C. Zhu, Direct observation of catch bonds involving cell-adhesion molecules, Nature423, 190 (2003)
2003
-
[39]
D. L. Huang, N. A. Bax, C. D. Buckley, W. I. Weis, and A. R. Dunn, Vinculin forms a directionally asymmetric 14 catch bond with F-actin, Science357, 703 (2017)
2017
-
[40]
V. C. Luca, B. C. Kim, C. Ge, S. Kakuda, D. Wu, M. Roein-Peikar, R. S. Haltiwanger, C. Zhu, T. Ha, and K. C. Garcia, Notch-jagged complex structure implicates a catch bond in tuning ligand sensitivity, Science355, 1320 (2017)
2017
-
[41]
Choi and C
H.-K. Choi and C. Zhu, Catch bonds in immunology, An- nual Review of Immunology43(2025)
2025
-
[42]
S. M. Block, Kinesin motor mechanics: Binding, step- ping, tracking, gating and limping, Biophys. J.92, 2986 (2007)
2007
-
[43]
Svoboda and S
K. Svoboda and S. M. Block, Force and velocity measured for single kinesin molecules, Cell77, 773 (1994)
1994
-
[44]
Svoboda and S
K. Svoboda and S. M. Block, Biological applications of optical forces, Annual review of biophysics and biomolec- ular structure23, 247 (1994)
1994
-
[45]
E. L. Holzbaur and Y. E. Goldman, Coordination of molecular motors: from in vitro assays to intracellular dynamics, Current opinion in cell biology22, 4 (2010)
2010
-
[46]
Isojima, R
H. Isojima, R. Iino, Y. Niitani, H. Noji, and M. Tomishige, Direct observation of intermediate states during the stepping motion of kinesin-1, Nature chemical biology12, 290 (2016)
2016
-
[47]
J.-K. Ryu, A. J. Katan, E. O. van der Sluis, T. Wisse, R. de Groot, C. H. Haering, and C. Dekker, The con- densin holocomplex cycles dynamically between open and collapsed states, Nature structural & molecular biology 27, 1134 (2020)
2020
-
[48]
Yildiz, M
A. Yildiz, M. Tomishige, R. D. Vale, and P. R. Selvin, Kinesin walks hand-over-hand, Science303, 676 (2004)
2004
-
[49]
A. B. Kolomeisky and M. E. Fisher, A simple kinetic model describes the processivity of myosin-V, Biophysi- cal journal84, 1642 (2003)
2003
-
[50]
Kodera, D
N. Kodera, D. Yamamoto, R. Ishikawa, and T. Ando, Video imaging of walking myosin v by high-speed atomic force microscopy, Nature468, 72 (2010)
2010
-
[51]
Hinczewski, R
M. Hinczewski, R. Tehver, and D. Thirumalai, Design principles governing the motility of yosin V, Proceedings of the National Academy of Sciences110, E4059 (2013)
2013
-
[52]
G. I. Bell, Models for the specific adhesion of cells to cells, Science200, 618 (1978)
1978
-
[53]
T. R. Strick, T. Kawaguchi, and T. Hirano, Real-time de- tection of single-molecule DNA compaction by condensin I, Current biology14, 874 (2004)
2004
-
[54]
D. T. Gillespie, Exact stochastic simulation of coupled chemical reactions, The journal of physical chemistry81, 2340 (1977)
1977
-
[55]
H. T. Vu, S. Chakrabarti, M. Hinczewski, and D. Thiru- malai, Discrete step sizes of molecular motors lead to bimodal non-gaussian velocity distributions under force, Physical review letters117, 078101 (2016)
2016
-
[56]
Dembo, Lectures on mathematics in the life sciences, some mathematical problems in biology, American Math- ematical Society, Providence, RI51, 51 (1994)
M. Dembo, Lectures on mathematics in the life sciences, some mathematical problems in biology, American Math- ematical Society, Providence, RI51, 51 (1994)
1994
-
[57]
Chakrabarti, M
S. Chakrabarti, M. Hinczewski, and D. Thirumalai, Phe- nomenological and microscopic theories for catch bonds, Journal of structural biology197, 50 (2017)
2017
-
[58]
Harder, A.-K
A. Harder, A.-K. M¨ oller, F. Milz, P. Neuhaus, V. Wal- horn, T. Dierks, and D. Anselmetti, Catch bond interac- tion between cell-surface sulfatase sulf1 and glycosamino- glycans, Biophysical journal108, 1709 (2015)
2015
-
[60]
Chakrabarti, M
S. Chakrabarti, M. Hinczewski, and D. Thiru- malai, Plasticity of hydrogen bond networks regulates mechanochemistry of cell adhesion complexes, Proceed- ings of the National Academy of Sciences111, 9048 (2014)
2014
-
[61]
C. O. Barkan and R. F. Bruinsma, Topology of molecular deformations induces triphasic catch bonding in selectin– ligand bonds, Proceedings of the National Academy of Sciences121, e2315866121 (2024)
2024
-
[62]
C. M. Johnson, J. D. Fenn, A. Brown, and P. Jung, Dy- namic catch-bonding generates the large stall forces of cytoplasmic dynein, Physical biology17, 046004 (2020)
2020
-
[63]
Pobegalov, L.-Y
G. Pobegalov, L.-Y. Chu, J.-M. Peters, and M. I. Molodtsov, Single cohesin molecules generate force by two distinct mechanisms, Nature Communications14, 3946 (2023)
2023
-
[64]
Ha and D
B.-Y. Ha and D. Thirumalai, A mean-field model for semiflexible chains, The Journal of chemical physics103, 9408 (1995)
1995
-
[65]
A. L. Nord, E. Gachon, R. Perez-Carrasco, J. A. Nirody, A. Barducci, R. M. Berry, and F. Pedaci, Catch bond drives stator mechanosensitivity in the bacterial flagellar motor, Proceedings of the National Academy of Sciences 114, 12952 (2017)
2017
-
[66]
M. Doi, S. F. Edwards, and S. F. Edwards,The theory of polymer dynamics, Vol. 73 (oxford university press, 1988)
1988
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.