Recognition: no theorem link
Dynamical mechanisms of flexible phase-locking in cortical theta oscillators
Pith reviewed 2026-05-11 02:21 UTC · model grok-4.3
The pith
Interactions between slow and superslow inhibitory currents expand the entrainment range of cortical theta oscillators through delayed Hopf phenomena.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
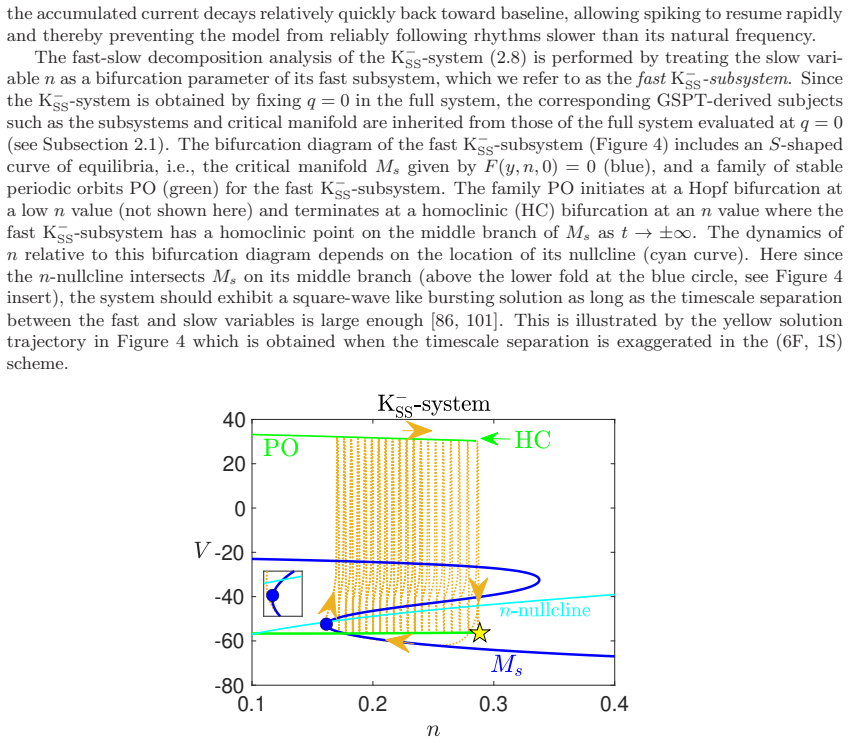
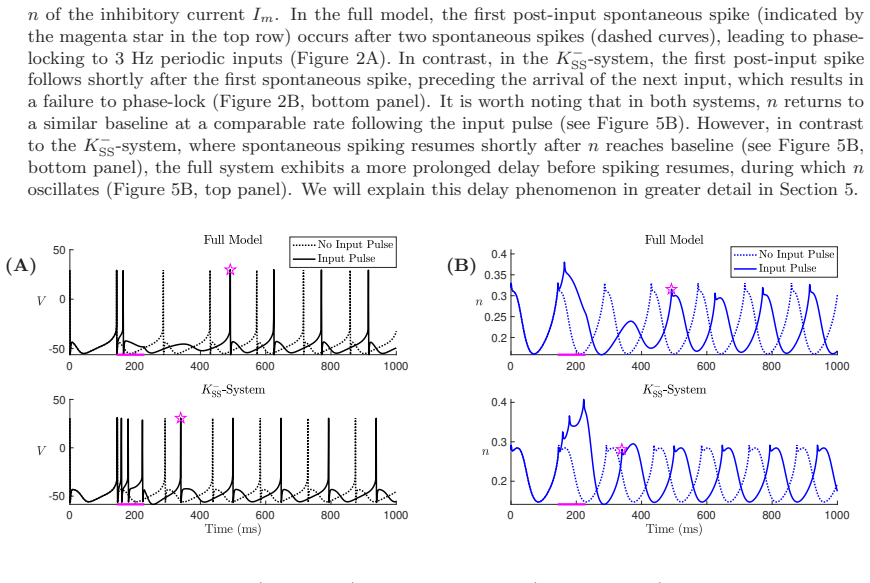
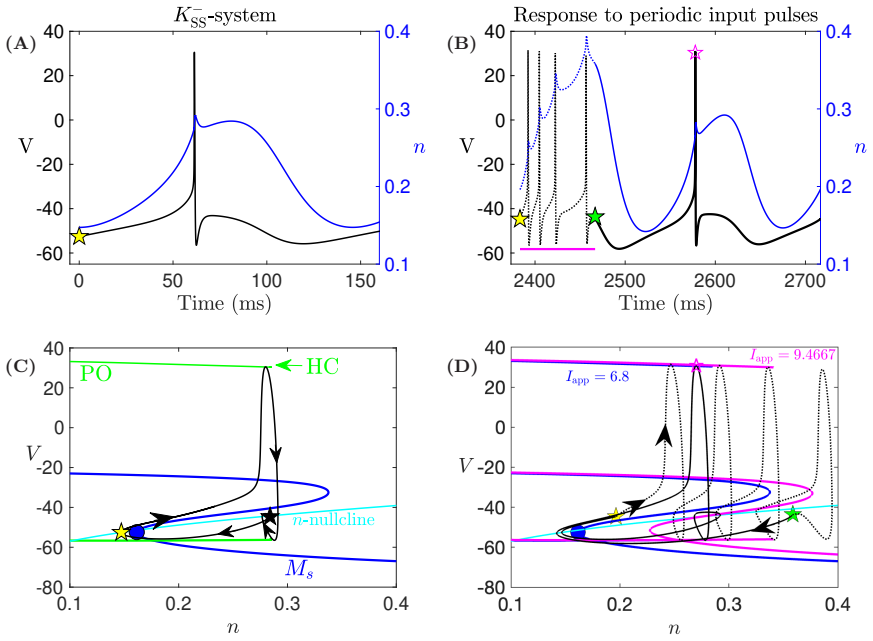
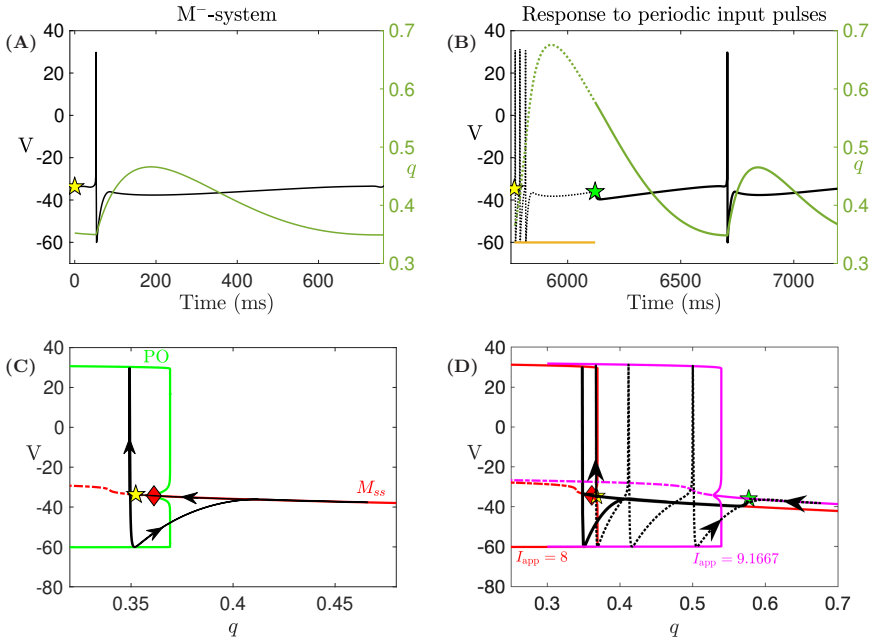
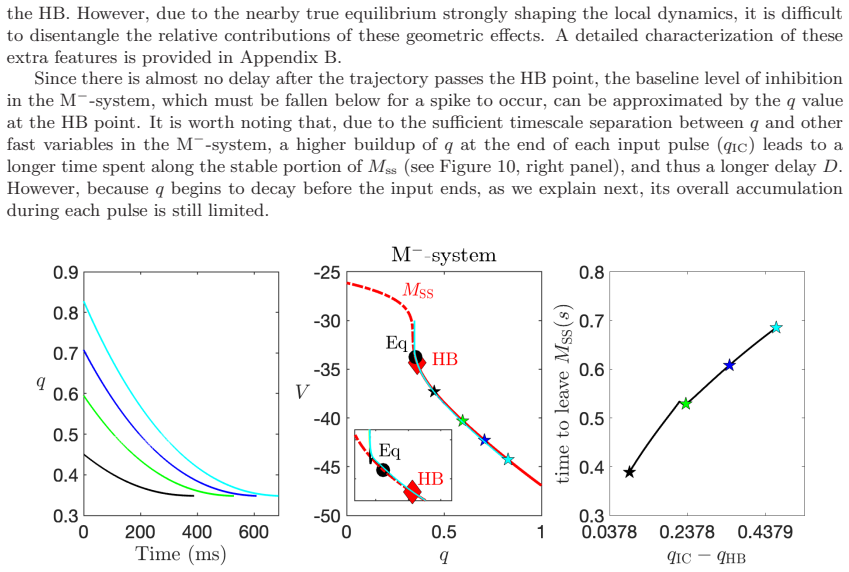
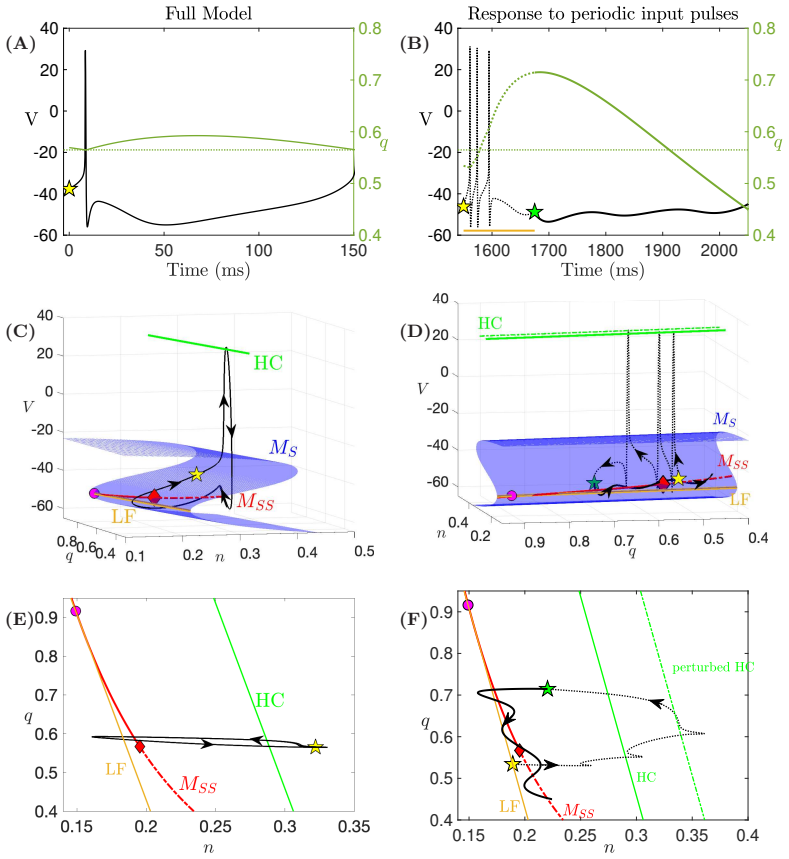
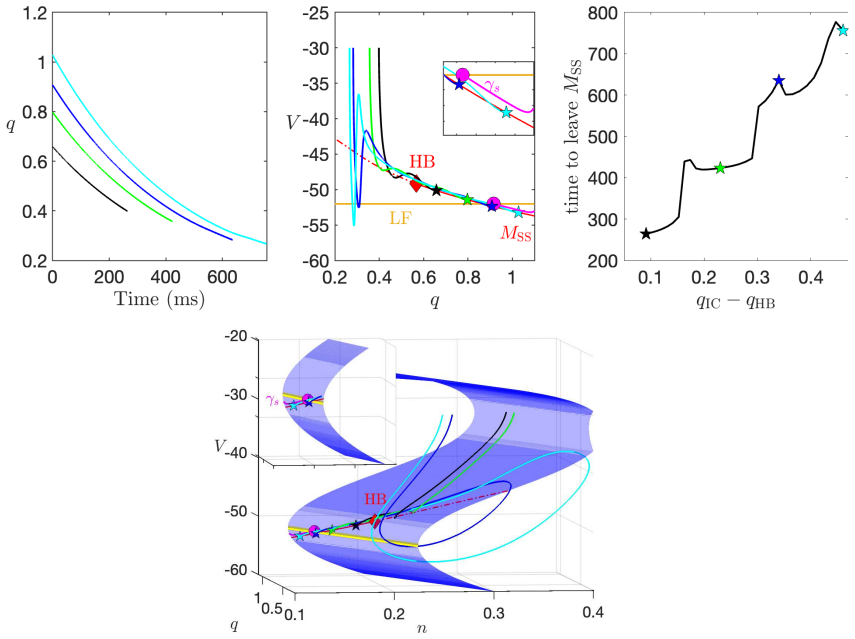
In a biophysically grounded model of cortical theta oscillators, the interaction between the theta-timescale inhibitory current I_m and the superslow delta-timescale potassium current I_K_SS produces a three-timescale dynamical structure. This structure generates pronounced post-input recovery delays associated with a delayed Hopf bifurcation. The superslow current has minimal impact on the unforced oscillation but is essential for expanding the phase-locking range under external input, while the intermediate current further prolongs the recovery along the superslow manifold.
What carries the argument
the delayed Hopf bifurcation (DHB) arising from interactions between slow and superslow inhibitory currents in a three-timescale system
If this is right
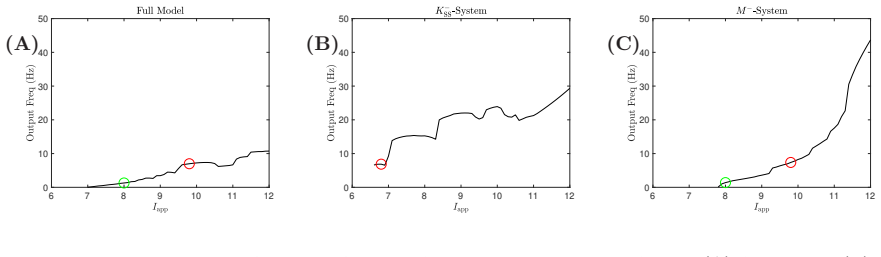
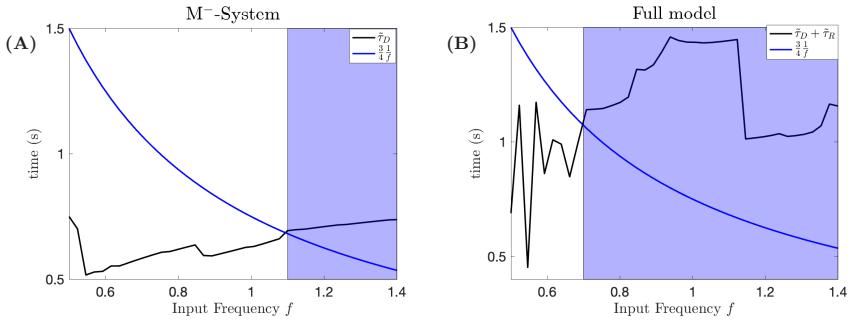
- The entrainment frequency range is substantially larger when both currents are present.
- Removing the superslow current reduces the oscillator's ability to lock to slower inputs.
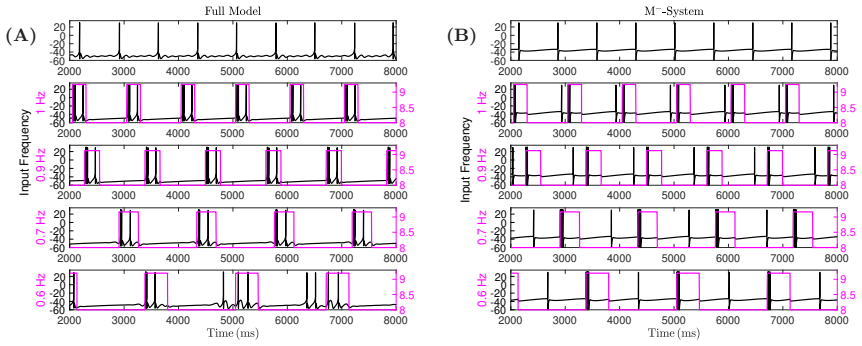
- The mechanism is recruited specifically by external forcing rather than being active in spontaneous oscillations.
- Coordination of multi-timescale inhibitory currents supports flexible phase-locking in cortical networks.
Where Pith is reading between the lines
- This could allow auditory cortex to track speech at varying speaking rates without changing the oscillator's intrinsic properties.
- Similar delayed recovery mechanisms might operate in other neural oscillators that need to synchronize to irregular or slow inputs.
- Experimental tests could involve blocking the superslow potassium current and measuring changes in entrainment flexibility in cortical slices.
Load-bearing premise
The model currents I_m and I_K_SS and the delayed Hopf bifurcation accurately represent the key dynamics responsible for flexible phase-locking in real cortical neurons.
What would settle it
Recording from cortical neurons during rhythmic input stimulation and finding no evidence of prolonged recovery delays linked to superslow inhibitory processes, or showing that the model's predicted entrainment range does not match observed neural synchronization.
Figures





read the original abstract
Oscillatory activity in auditory cortex is thought to play a central role in auditory and speech processing by synchronizing neural rhythms to external acoustic features of the speech stream. To support this function, cortical oscillators must flexibly phase-lock to inputs spanning a wide range of timescales, including rhythms substantially slower than their intrinsic frequency. Here we identify a general dynamical mechanism by which intrinsic inhibitory currents operating on multiple timescales enable such flexible phase-locking. Using tools from dynamical systems theory, we show that interactions between slow and superslow inhibitory processes generate prolonged post-input recovery delays through delayed Hopf phenomena, thereby substantially expanding the frequency range over which entrainment can occur. We demonstrate this mechanisms in a biophysically grounded cortical theta oscillator model for speech segmentation. Specifically, we show that both a theta-timescale (4-8 Hz) inhibitory current $I_m$ and a slower delta-timescale (1-4 Hz) inhibitory potassium current $I_{\rm K_{SS}}$ are crucial for entrainment flexibility. Their interaction creates a three-timescale structure that gives rise to pronounced delay phenomena associated with a delayed Hopf bifurcation (DHB). Interestingly, the superslow $I_{\rm K_{SS}}$ and the associated DHB play little role in the unforced oscillatory dynamics, but are recruited to support phase locking under external forcing. Moreover, the intermediate-timescale current $I_m$, rather than being redundant, further expands the phase-locking range by prolonging delayed recovery along the superslow manifold. Together, these results suggest that coordination among intrinsic inhibitory currents operating on multiple timescales may represent a key mechanism supporting flexible phase locking to rhythmic inputs in the brain.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that in a biophysically grounded cortical theta oscillator model, interactions between a theta-timescale inhibitory current I_m and a superslow delta-timescale potassium current I_K_SS create a three-timescale dynamical structure. This structure produces prolonged post-input recovery delays via delayed Hopf bifurcation (DHB) phenomena, substantially expanding the frequency range for flexible phase-locking to external rhythmic inputs, with relevance to auditory cortex and speech segmentation. The superslow current is said to play little role in unforced dynamics but is recruited under forcing, while I_m further expands the locking range by prolonging recovery along the superslow manifold.
Significance. If the mechanism holds, the work provides a concrete dynamical-systems account of how multi-timescale inhibition supports entrainment beyond an oscillator's intrinsic frequency, which could be relevant for models of auditory and speech processing. The emphasis on delayed Hopf phenomena as a general principle for flexible phase-locking, identified through analysis of a conductance-based model, is a potential strength if supported by targeted verification.
major comments (2)
- [Results on forced dynamics and DHB analysis] The attribution of expanded entrainment specifically to the delayed Hopf bifurcation arising from I_m / I_K_SS interaction is load-bearing for the central claim but lacks direct causal verification. The manuscript reports forward simulations showing recruitment of I_K_SS under forcing and prolongation by I_m, yet does not include ablation experiments (e.g., setting the conductance of I_K_SS to zero while retaining I_m) to test whether the entrainment bandwidth collapses, nor manifold continuation past the putative DHB point to quantify the delay scaling. Without these, the necessity of the DHB mechanism versus effects of the slow current alone remains an interpretation.
- [Dynamical systems analysis and entrainment simulations] The three-timescale structure is presented as enabling pronounced delay phenomena, but the quantitative mapping from the DHB to the observed expansion in locking range (e.g., specific frequency bounds tested and effect sizes) is not tied to explicit bifurcation diagrams or delay estimates in the reported figures. This weakens the link between the dynamical mechanism and the functional claim of 'substantially expanding' the entrainment range.
minor comments (2)
- [Model description] Notation for the currents (I_m and I_{K_SS}) is introduced clearly in the abstract but should be cross-referenced with explicit equations in the model section for readers unfamiliar with the specific conductances.
- [Abstract and unforced dynamics] The abstract states that I_K_SS 'plays little role in the unforced dynamics'; a brief quantification of this (e.g., change in intrinsic frequency or stability when I_K_SS is removed) would strengthen the contrast with its role under forcing.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which highlight important opportunities to strengthen the causal evidence for the delayed Hopf bifurcation mechanism. We have revised the manuscript accordingly and address each major comment below.
read point-by-point responses
-
Referee: The attribution of expanded entrainment specifically to the delayed Hopf bifurcation arising from I_m / I_K_SS interaction is load-bearing for the central claim but lacks direct causal verification. The manuscript reports forward simulations showing recruitment of I_K_SS under forcing and prolongation by I_m, yet does not include ablation experiments (e.g., setting the conductance of I_K_SS to zero while retaining I_m) to test whether the entrainment bandwidth collapses, nor manifold continuation past the putative DHB point to quantify the delay scaling. Without these, the necessity of the DHB mechanism versus effects of the slow current alone remains an interpretation.
Authors: We agree that ablation experiments and manifold continuation would provide stronger causal support. In the revised manuscript we have added simulations in which the conductance of I_K_SS is set to zero while I_m is retained; these show a clear collapse of the entrainment bandwidth, indicating that the superslow current is necessary for the expanded range. We have also performed and included numerical continuation along the superslow manifold past the DHB, which quantifies the delay scaling and demonstrates that the prolongation arises specifically from the delayed Hopf phenomenon rather than from the slow current in isolation. revision: yes
-
Referee: The three-timescale structure is presented as enabling pronounced delay phenomena, but the quantitative mapping from the DHB to the observed expansion in locking range (e.g., specific frequency bounds tested and effect sizes) is not tied to explicit bifurcation diagrams or delay estimates in the reported figures. This weakens the link between the dynamical mechanism and the functional claim of 'substantially expanding' the entrainment range.
Authors: We accept that the original figures did not make the quantitative connection explicit enough. The revised manuscript now includes bifurcation diagrams that locate the DHB within the three-timescale system and directly relate it to the frequency bounds used in the entrainment simulations. We have also added explicit delay estimates obtained from the DHB analysis, showing how the mechanism produces the observed expansion in locking range. These additions tie the dynamical structure to the functional results more rigorously. revision: yes
Circularity Check
No circularity: derivation rests on independent dynamical analysis of the model
full rationale
The paper constructs a biophysically grounded theta oscillator model incorporating I_m and I_K_SS, then applies standard dynamical systems tools (phase-plane analysis, manifold reduction, bifurcation tracking) to demonstrate that their interaction produces delayed Hopf phenomena and expanded entrainment range. The central claim is obtained by forward integration and continuation of the model's ODEs under forcing; it is not obtained by fitting parameters to the target entrainment bandwidth, redefining the output as input, or invoking self-citations whose content is presupposed. The statements that I_K_SS is recruited only under forcing and that I_m prolongs recovery along the superslow manifold are direct consequences of the three-timescale structure and the location of the DHB, not tautological restatements. No load-bearing step reduces to a fitted input or to a prior result by the same authors that itself lacks independent verification.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Cortical theta oscillators can be modeled using biophysically grounded equations incorporating theta-timescale inhibitory current I_m and delta-timescale potassium current I_K_SS
Reference graph
Works this paper leans on
-
[1]
Srisairam Achuthan and Carmen C Canavier. Phase-resett ing curves determine synchroniza- tion, phase locking, and clustering in networks of neural os cillators. Journal of Neuroscience , 29(16):5218–5233, 2009
work page 2009
-
[2]
Synchroniz ation of strongly coupled excitatory neurons: relating network behavior to biophysics
Corey D Acker, Nancy Kopell, and John A White. Synchroniz ation of strongly coupled excitatory neurons: relating network behavior to biophysics. Journal of computational neuroscience , 15(1):71– 90, 2003. 24
work page 2003
- [3]
-
[4]
The brain’s best kept secret is its degenerate structure
Larissa Albantakis, Christophe Bernard, Naama Brenner , Eve Marder, and Rishikesh Narayanan. The brain’s best kept secret is its degenerate structure. Journal of Neuroscience , 44(40), 2024
work page 2024
- [5]
-
[6]
S. M. Baer, T. Erneux, and J. Rinzel. The slow passage thro ugh a Hopf bifurcation: Delay, memory effects, and resonance. SIAM J. Appl. Math. , 49(1):55–71, 1989
work page 1989
-
[7]
H. Baldemir, D. Avitabile, and K. Tsaneva-Atanasova. Ps eudo-plateau bursting and mixed-mode oscillations in a model of developing inner hair cells. Commun Nonlinear Sci Numer Simulat , 80:104979, 2020
work page 2020
-
[8]
S. Battaglin and M. G. Pedersen. Geometric analysis of mi xed-mode oscillations in a model of electrical activity in human beta-cells. Nonlinear Dyn. , 104(4):4445–4457, 2021
work page 2021
-
[9]
¨Uber das elektroenkephalogramm des menschen
Hans Berger. ¨Uber das elektroenkephalogramm des menschen. Archiv f¨ ur psychiatrie und ner- venkrankheiten, 87(1):527–570, 1929
work page 1929
- [10]
-
[11]
Gy¨ orgy Buzs´ aki.Rhythms of the Brain . Oxford university press, 2006
work page 2006
-
[12]
The origin of extracellular fields and currents—eeg, ecog, lfp and spikes
Gy¨ orgy Buzs´ aki, Costas A Anastassiou, and Christof K och. The origin of extracellular fields and currents—eeg, ecog, lfp and spikes. Nature reviews neuroscience , 13(6):407–420, 2012
work page 2012
-
[13]
Neuronal oscillat ions in cortical networks
Gyorgy Buzsaki and Andreas Draguhn. Neuronal oscillat ions in cortical networks. science, 304(5679):1926–1929, 2004
work page 1926
-
[14]
Pulse couple d oscillators and the phase resetting curve
Carmen C Canavier and Srisairam Achuthan. Pulse couple d oscillators and the phase resetting curve. Mathematical biosciences, 226(2):77–96, 2010
work page 2010
-
[15]
Jonathan Cannon and Nancy Kopell. The leaky oscillator : Properties of inhibition-based rhythms revealed through the singular phase response curve. SIAM Journal on Applied Dynamical Systems , 14(4):1930–1977, 2015
work page 1930
-
[16]
L. M. Carracedo, H. Kjeldsen, L. Cunnington, A. Jenkins , I. Schofield, and et al. M.O. Cunningham, M. O. A neocortical delta rhythm facilitates reciprocal int erlaminar interactions via nested theta rhythms. Journal of Neuroscience , 33(26):10750–10761, 2013
work page 2013
-
[17]
P hase-amplitude response functions for transient-state stimuli
Oriol Castej´ on, Antoni Guillamon, and Gemma Huguet. P hase-amplitude response functions for transient-state stimuli. The Journal of Mathematical Neuroscience , 3(1):13, 2013
work page 2013
-
[18]
The natural statistics of audiovisual speech
Chandramouli Chandrasekaran, Andrea Trubanova, S´ ebastien Stillittano, Alice Caplier, and Asif A Ghazanfar. The natural statistics of audiovisual speech. PLoS computational biology, 5(7):e1000436, 2009
work page 2009
-
[19]
G. A. Chumakov, N. A. Chumakova, and E. A. Lashina. Model ing the complex dynamics of heterogeneous catalytic reactions with fast, intermediat e, and slow variables. Chem. Eng. J. , 282:11–19, 2015
work page 2015
-
[20]
Fun ctional phase response curves: a method for understanding synchronization of adapting neurons
Jianxia Cui, Carmen C Canavier, and Robert J Butera. Fun ctional phase response curves: a method for understanding synchronization of adapting neurons. Journal of Neurophysiology , 102(1):387– 398, 2009
work page 2009
-
[21]
R. Curtu. Singular Hopf bifurcation and mixed-mode osc illations in a two-cell inhibitory neural network. Phys. D: Nonlinear Phenom. , 239(9):504–514, 2010
work page 2010
-
[22]
R. Curtu and J. Rubin. Interaction of canard and singula r Hopf mechanisms in a neural model. SIAM J. Appl. Dyn. Syst. , 10(4):1443–1479, 2011
work page 2011
-
[23]
P. De Maesschalck, E. Kutafina, and N. Popovi´ c. Three ti me-scales in an extended Bonhoeffer–van der Pol oscillator. J. Dyn. Differ. Equ. , 26:955–987, 2014. 25
work page 2014
-
[24]
P. De Maesschalck, E. Kutafina, and N. Popovi´ c. Sector-delayed-Hopf-type mixed-mode oscillations in a prototypical three-time-scale model. Appl. Math. Comput. , 273:337–352, 2016
work page 2016
-
[25]
M. Desroches, J. Guckenheimer, B. Krauskopf, C. Kuehn, H. M. Osinga, and M. Wechselberger. Mixed-mode oscillations with multiple time scales. SIAM Rev. , 54(2):211–288, 2012
work page 2012
-
[26]
M. Desroches and V. Kirk. Spike-adding in a canonical th ree-time-scale model: superslow explosion and folded-saddle canards. SIAM J. Appl. Dyn. Syst. , 17(3):1989–2017, 2018
work page 1989
-
[27]
Temporal modulations in speech and music
Nai Ding, Aniruddh D Patel, Lin Chen, Henry Butler, Chen g Luo, and David Poeppel. Temporal modulations in speech and music. Neuroscience & Biobehavioral Reviews , 81:181–187, 2017
work page 2017
- [28]
-
[29]
Degeneracy and comp lexity in biological systems
Gerald M Edelman and Joseph A Gally. Degeneracy and comp lexity in biological systems. Pro- ceedings of the national academy of sciences , 98(24):13763–13768, 2001
work page 2001
-
[30]
D elays and advances in the onset of instability in the shishkova equation
Hans Engler, Hans Kaper, Tasso Kaper, and Theodore Vo. D elays and advances in the onset of instability in the shishkova equation. Quarterly of Applied Mathematics , 2025
work page 2025
-
[31]
B. Ermentrout. Type i membranes, phase resetting curve s, and synchrony. Neural Comput., 8:979– 1001, 1996
work page 1996
-
[32]
Type i membranes, phase resetting cur ves, and synchrony
Bard Ermentrout. Type i membranes, phase resetting cur ves, and synchrony. Neural computation, 8(5):979–1001, 1996
work page 1996
-
[33]
Bard Ermentrout, Matthew Pascal, and Boris Gutkin. The effects of spike frequency adaptation and negative feedback on the synchronization of neural osci llators. Neural computation, 13(6):1285– 1310, 2001
work page 2001
-
[34]
Mathematical foundations of neuroscience , volume 35
Bard Ermentrout and David Hillel Terman. Mathematical foundations of neuroscience , volume 35. Springer, 2010
work page 2010
-
[35]
n: m phase-locking of weakly coupled oscillators
G Bard Ermentrout. n: m phase-locking of weakly coupled oscillators. Journal of Mathematical Biology, 12(3):327–342, 1981
work page 1981
-
[36]
Multiple pulse inte ractions and averaging in systems of coupled neural oscillators
G Bard Ermentrout and Nancy Kopell. Multiple pulse inte ractions and averaging in systems of coupled neural oscillators. Journal of Mathematical Biology , 29(3):195–217, 1991
work page 1991
- [37]
-
[38]
O. Ghitza. Behavioral evidence for the role of cortical theta oscillations in determining auditory channel capacity for speech. Frontiers in psychology , 5:652, 2014
work page 2014
-
[39]
O. Ghitza and S. Greenberg. On the possible role of brain rhythms in speech perception: intel- ligibility of time-compressed speech with periodic and ape riodic insertions of silence. Phonetica, 66(1-2):113–126, 2009
work page 2009
-
[40]
Oded Ghitza. Linking speech perception and neurophysi ology: speech decoding guided by cascaded oscillators locked to the input rhythm. Frontiers in psychology , 2:130, 2011
work page 2011
-
[41]
Oded Ghitza. On the role of theta-driven syllabic parsi ng in decoding speech: intelligibility of speech with a manipulated modulation spectrum. Frontiers in psychology , 3:238, 2012
work page 2012
-
[42]
Gorsev Giilmen Yenera’c’d’e. Brain’s alpha, beta, gamma, delta, and theta oscillationsin neuropsy- chiatric diseases: proposal for biomarker strategies
-
[43]
Ion channel degene racy, variability, and covariation in neuron and circuit resilience
Jean-Marc Goaillard and Eve Marder. Ion channel degene racy, variability, and covariation in neuron and circuit resilience. Annual review of neuroscience , 44(1):335–357, 2021
work page 2021
-
[44]
Speaking in shorthand–a syllable-c entric perspective for understanding pronun- ciation variation
Steven Greenberg. Speaking in shorthand–a syllable-c entric perspective for understanding pronun- ciation variation. Speech Communication, 29(2-4):159–176, 1999
work page 1999
-
[45]
J. Guckenheimer and A. R. Willms. Asymptotic analysis o f subcritical Hopf-homoclinic bifurcation. Phys. D: Nonlinear Phenom. , 139(3-4):195–216, 2000
work page 2000
-
[46]
Y. Gutfreund, Y. Yarom, and I. Segev. Subthreshold osci llations and resonant frequency in guinea- pig cortical neurons: physiology and modelling. The Journal of physiology , 483(3):621–640, 1985. 26
work page 1985
- [47]
- [48]
-
[49]
M. G. Hayes, T. J. Kaper, P. Szmolyan, and M. Wechselberg er. Geometric desingularization of degenerate singularities in the presence of fast rotation: A new proof of known results for slow passage through Hopf bifurcations. Indag. Math. , 27(5):1184–1203, 2016
work page 2016
-
[50]
Sevada Hovsepyan, Itsaso Olasagasti, and Anne-Lise Gi raud. Combining predictive coding and neural oscillations enables online syllable recognition i n natural speech. Nature communications, 11(1):3117, 2020
work page 2020
-
[51]
J. L. Hudson, M. Hart, and D. Marinko. An experimental st udy of multiple peak periodic and nonperiodic oscillations in the Belousov–Zhabotinskii re action. J. Chem. Phys. , 71(4):1601–1606, 1979
work page 1979
- [52]
-
[53]
Speech encoding by coupled cortical theta and gamma oscilla tions
Alexandre Hyafil, Lorenzo Fontolan, Claire Kabdebon, B oris Gutkin, and Anne-Lise Giraud. Speech encoding by coupled cortical theta and gamma oscilla tions. elife, 4:e06213, 2015
work page 2015
- [54]
-
[55]
P. Kaklamanos and N. Popovi´ c. Complex oscillatory dyn amics in a three-timescale El Ni˜ no South- ern Oscillation model. Phys. D: Nonlinear Phenom. , 449:133740, 2023
work page 2023
-
[56]
P. Kaklamanos, N. Popovi´ c, and K. U. Kristiansen. Bifu rcations of mixed-mode oscillations in three-timescale systems: An extended prototypical exampl e. Chaos: An Interdisciplinary Journal of Nonlinear Science , 32(1):013108, 2022
work page 2022
-
[57]
P. Kaklamanos, N. Popovi´ c, and K. U. Kristiansen. Geom etric singular perturbation analysis of the multiple-timescale Hodgkin-Huxley equations. SIAM J. Appl. Dyn. Syst. , 22(3):1552–1589, 2023
work page 2023
- [58]
- [59]
-
[60]
Phase response func- tion for oscillators with strong forcing or coupling
Vladimir Klinshov, Serhiy Yanchuk, Artur Stephan, and Vladimir Nekorkin. Phase response func- tion for oscillators with strong forcing or coupling. Europhysics Letters, 118(5):50006, 2017
work page 2017
-
[61]
Mechanisms of phase -locking and frequency control in pairs of coupled neural oscillators
Nancy Kopell and G Bard Ermentrout. Mechanisms of phase -locking and frequency control in pairs of coupled neural oscillators. Handbook of dynamical systems , 2:3–54, 2002
work page 2002
- [62]
- [63]
- [64]
-
[65]
M. Krupa and M. Wechselberger. Local analysis near a fol ded saddle-node singularity. J. Differ. Equ., 248(12):2841–2888, 2010
work page 2010
-
[66]
P. K¨ ugler, A. H. Erhardt, and M. A. K. Bulelzai. Early af terdepolarizations in cardiac action potentials as mixed mode oscillations due to a folded node si ngularity. PLoS One, 13(12):e0209498, 2018. 27
work page 2018
-
[67]
Peter Lakatos, Ankoor S Shah, Kevin H Knuth, Istvan Ulbe rt, George Karmos, and Charles E Schroeder. An oscillatory hierarchy controlling neuronal excitability and stimulus processing in the auditory cortex. Journal of neurophysiology , 94(3):1904–1911, 2005
work page 1904
- [68]
-
[69]
Phase resetting reduce s theta–gamma rhythmic interaction to a one-dimensional map
Paola Malerba and Nancy Kopell. Phase resetting reduce s theta–gamma rhythmic interaction to a one-dimensional map. Journal of mathematical biology , 66(7):1361–1386, 2013
work page 2013
-
[70]
Synchronizatio n of pulse-coupled biological oscillators
Renato E Mirollo and Steven H Strogatz. Synchronizatio n of pulse-coupled biological oscillators. SIAM Journal on Applied Mathematics , 50(6):1645–1662, 1990
work page 1990
-
[71]
C. Morris and H. Lecar. Voltage oscillations in the barn acle giant muscle fiber. Biophys. J. , 35(1):193–213, 1981
work page 1981
-
[72]
P. Nan, Y. Wang, V. Kirk, and J. E. Rubin. Understanding a nd distinguishing three-time-scale oscillations: Case study in a coupled Morris-Lecar system. SIAM J. Appl. Dyn. Syst. , 14(3):1518– 1557, 2015
work page 2015
- [73]
- [74]
-
[75]
The temporal regulation of speech
John J Ohala. The temporal regulation of speech. Auditory analysis and perception of speech , pages 431–453, 1975
work page 1975
-
[76]
The utility of phase models in studying neural synchronization
Youngmin Park, Stewart Heitmann, and G Bard Ermentrout . The utility of phase models in studying neural synchronization. Computational models of brain and behavior , pages 493–504, 2017
work page 2017
-
[77]
E. Pavlidis, F. Campillo, A. Goldbeter, and M. Desroche s. Multiple-timescale dynamics, mixed mode oscillations and mixed affective states in a model of bip olar disorder. Cognitive Neurodynam- ics, 2022
work page 2022
-
[78]
Global ph ase-amplitude description of oscillatory dy- namics via the parameterization method
Alberto P´ erez-Cervera, Gemma Huguet, et al. Global ph ase-amplitude description of oscillatory dy- namics via the parameterization method. Chaos: an interdisciplinary journal of nonlinear science , 30(8), 2020
work page 2020
-
[79]
Phase-locked states in oscillating neural networks and their role in neural communication
Alberto P´ erez-Cervera, Tere M Seara, and Gemma Huguet . Phase-locked states in oscillating neural networks and their role in neural communication. Communications in Nonlinear Science and Numerical Simulation , 80:104992, 2020
work page 2020
-
[80]
C. Perryman and S. Wieczorek. Adapting to a changing env ironment: non-obvious thresholds in multi-scale systems. Proc. Math. Phys. Eng. Sci. , 470(2170):20140226, 2014
work page 2014
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.