Recognition: no theorem link
Logarithmic scaling of selective sweep curves: from tents to houses
Pith reviewed 2026-05-12 04:12 UTC · model grok-4.3
The pith
Logarithmic scaling turns selective sweep curves into tents for strong selection and houses for moderate selection in the large population limit.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
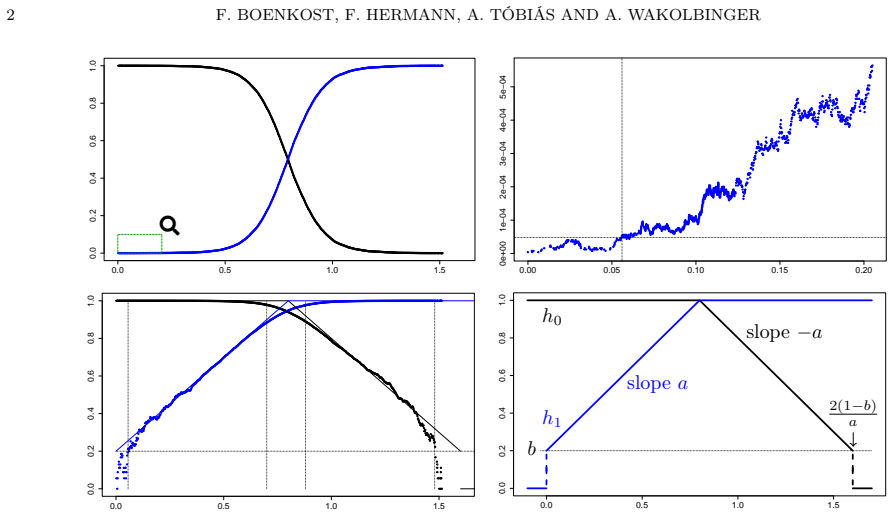
In the Moran model, logarithmic scaling of selective sweep curves produces convergence to a tent shape under strong selection and to a house shape under moderate selection in the large population limit; the convergence is uniform on the roof (linear growth and decline phases) and holds in the Skorokhod M1 topology on the walls (near the jumps).
What carries the argument
Logarithmic scaling of mutant frequency and time, which transforms logistic sweep curves into piecewise-linear tent or house shapes whose convergence regularity is measured by uniform topology on linear segments and Skorokhod M1 topology near jumps.
If this is right
- The result supplies the technical foundation needed to extend Poissonian interacting trajectory descriptions of clonal interference from strong selection to moderate selection.
- Approximations of fixation times and frequencies in large populations can now be made rigorous by matching observed trajectories to the tent or house limit shapes.
- The distinction between tent and house geometries quantifies how selection strength controls the presence or absence of abrupt frequency jumps at the beginning and end of a sweep.
Where Pith is reading between the lines
- The same scaling and topology framework may apply to other standard models such as the Wright-Fisher process, offering a route to compare sweep behavior across common population-genetic simulators.
- Genomic data showing frequency trajectories could be matched against the tent or house shapes to estimate whether selection was strong or moderate without assuming a specific model beyond the large-population limit.
- The house shape's jumps suggest that moderate selection produces detectable signatures in allele-frequency time series that strong selection lacks, which could be tested in forward simulations of finite but growing populations.
Load-bearing premise
The population evolves according to the Moran model and the large-population limit is taken with either strong or moderate selection.
What would settle it
A direct simulation of the Moran process in the large-population regime that shows the scaled curves deviate from uniform convergence on the linear phases or fail to match the Skorokhod M1 distance near the jumps would falsify the claimed regularity.
Figures

read the original abstract
One of the classical results of mathematical population genetics states that the frequency of a beneficial mutant's offspring, on its way to fixation in a large population, looks like a logistic curve. A logarithmic scaling (both in height and time) of these selective sweep curves leads (in the case of strong selection) to a tent-like shape in the large population limit: First the logarithmic frequency of the mutant increases linearly from 0 to 1, then that of the former resident decreases from 1 to 0. For moderate selection the logarithmic frequencies develop (in the large population limit) a jump at the beginning/the end of the sweep, which takes the shape of the tent into that of a house. Our main result (proved for the Moran model) assesses the regularity of this convergence in the large population limit: It is uniform in the house's roof (phases of linear growth and decline) and "Skorokhod $M_1$" in the house's walls (closely around the jumps). Apart from interest in its own right, we anticipate that this result and the proof techniques will be instrumental for extending the description of clonal interference by Poissonian interacting trajectories (as it was done in Hermann et al. (2024) for strong selection) also to moderate selection.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proves a limit theorem for logarithmically scaled selective sweep curves in the Moran model. Under strong selection the curves converge to a tent shape in the large-population limit; under moderate selection they converge to a house shape with jumps. The main result establishes that this convergence is uniform on the linear phases (roof) and holds in the Skorokhod M1 topology near the jumps (walls). The proof is carried out explicitly for the Moran process and the techniques are presented as a foundation for extending clonal-interference descriptions to moderate selection.
Significance. The result supplies a precise regularity statement for a classical object in mathematical population genetics. Uniform convergence on the linear segments and M1 convergence at the discontinuities are the right topologies for the intended downstream applications to interacting trajectories. By working in the Moran model the authors obtain a fully rigorous proof whose methods can be reused; this is a concrete strength that supports the claim that the work will help extend Hermann et al. (2024) to moderate selection.
minor comments (4)
- §2: the informal description of the 'house' shape is clear, but the precise definition of the limiting process (including the location and size of the jumps) should be stated as a displayed equation before the main theorem is formulated.
- Theorem 3.2 (or equivalent): the statement of Skorokhod M1 convergence near the jumps would benefit from an explicit reference to the metric used and a one-sentence reminder why M1 is preferred over J1 in this setting.
- Proof of the uniform convergence on the roof (around Eq. (14)–(18)): the dependence of the uniformity constants on the selection coefficient s should be tracked explicitly so that the moderate-selection regime is visibly covered.
- Introduction, paragraph 3: a short sentence recalling the classical logistic-curve result (with citation) would help readers who are not already familiar with the unscaled sweep.
Simulated Author's Rebuttal
We thank the referee for the positive assessment of our manuscript on logarithmic scaling of selective sweep curves and for recommending minor revision. The report contains no major comments, so we have no points to address point-by-point. We will incorporate any minor suggestions that may be provided in the full report or during revision.
Circularity Check
Minor self-citation; central proof is independent
full rationale
The paper's core contribution is a mathematical theorem establishing uniform convergence on linear phases and Skorokhod M1 topology near jumps for logarithmically scaled selective sweep curves in the Moran model, under large-population limits with strong or moderate selection. This is derived via stochastic process analysis and does not involve parameter fitting, self-definitional constructs, or reductions by construction. A reference to Hermann et al. (2024) provides context for the strong-selection case and future extensions to clonal interference, but the present proof for moderate selection and regularity assessment stands independently without load-bearing reliance on the citation. No load-bearing steps in the derivation chain reduce to inputs by definition or fitting.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The Moran model accurately represents population dynamics for the selective sweep process.
Reference graph
Works this paper leans on
-
[1]
K. S. Alexander. Excursions and local limit theorems for B essel-like random walks. Electron.\ J.\ Probab., 16 0 (paper no.\ 1): 0 1--44, 2011
work page 2011
-
[2]
K. B. Athreya and P. E. Ney. Branching processes. Springer-Verlag Berlin, Mineola, NY, 1972
work page 1972
- [3]
-
[4]
A. D. Barbour, K. Hamza, H. Kaspi, and F. C. Klebaner. Escape from the boundary in M arkov population processes. Advances in Applied Probability, 47 0 (4): 0 1190--1211, 2015
work page 2015
-
[5]
N. H. Barton. The effect of hitch-hiking on neutral genealogies. Genetics Research, 72 0 (2): 0 123--133, 1998
work page 1998
-
[6]
N. Champagnat, S. Méléard, and V. C. Tran. Stochastic analysis of emergence of evolutionary cyclic behavior in population dynamics with transfer. Annals of Applied Probability, 31 0 (4): 0 1820--1867, 2021. ISSN 1742-5662
work page 2021
-
[7]
K. L. Chung, J. B. Walsh, et al. To reverse a M arkov process. Acta Math, 123 0 (1): 0 225--251, 1969
work page 1969
-
[8]
F. Cordero. The deterministic limit of the M oran model: a uniform central limit theorem. Markov Processes and Related Fields, 23: 0 313--324, 2017
work page 2017
-
[9]
C. Desmarais, E. Schertzer, and Zs. Talyig \'a s. K -branching random walk with noisy selection: large population limits and phase transitions. arXiv preprint arXiv:2509.26254, 2025
-
[10]
A. Etheridge, P. Pfaffelhuber, and A. Wakolbinger. An approximate sampling formula under genetic hitchhikng. Annals of Applied Probability, 16: 0 685--729, 2006
work page 2006
-
[11]
S. N. Ethier and Th. G. Kurtz. Markov processes: characterization and convergence. John Wiley & Sons, 2009
work page 2009
-
[12]
W. J. Ewens. Mathematical population genetics: theoretical introduction, volume 27. Springer, 2004
work page 2004
-
[13]
B. M. Hambly, G. Kersting, and A. E. Kyprianou. Law of the iterated logarithm for oscillating random walks conditioned to stay non-negative. Stochastic processes and their applications, 108 0 (2): 0 327--343, 2003
work page 2003
-
[14]
S. C. Harris, S. G. G. Johnston, and M. I. Roberts. The coalescent structure of continuous-time G alton-- W atson trees. Annals of Applied Probability, 30: 0 1368--1414, 2020
work page 2020
-
[15]
T. E. Harris. The theory of branching processes. Die Grundlehren der Mathematischen Wissenschaften, Bd. 119. Springer-Verlag, Berlin, 1963
work page 1963
-
[16]
F. Hermann, A. Gonz\'alez\, Casanova, R. S oares dos Santos , A. T\'obi\'as, and A. Wakolbinger. From clonal interference to P oissonian interacting trajectories. Annals of Applied Probability, 35 0 (4): 0 2823--2865, 2025
work page 2025
-
[17]
N. L. Kaplan, R. R. Hudson, and C. H. Langley. The hitch-hiking effect re-visited. Genetics, 123: 0 887--899, 1989
work page 1989
-
[18]
J. Kern. S korokhod topologies. Mathematische Semesterberichte, 71: 0 1--18, 2024
work page 2024
- [19]
-
[20]
J. W. Pitman. One-dimensional B rownian motion and the three-dimensional B essel process. Advances in Applied Probability, 6 0 (2): 0 223--224, 1974
work page 1974
-
[21]
A. V. Skorokhod. Limit theorems for stochastic processes. Theory of Probability & Its Applications, 1 0 (3): 0 261--290, 1956
work page 1956
-
[22]
J. M. Smith and J. Haigh. The hitch-hiking effect of a favourable gene. Genetics Research, 23 0 (1): 0 23--35, 1974
work page 1974
-
[23]
N. Udomchatpitak and J. Schweinsberg. The accumulation of beneficial mutations and convergence to a P oisson process. Stochastic Processes and their Applications, 183: 0 104578, 2025
work page 2025
-
[24]
W. Whitt. Stochastic-Process Limits. Springer Series in Operations Research and Financial Engineering (ORFE). Springer, 2006
work page 2006
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.