Recognition: no theorem link
Kin-ematic Exclusion in Active Matter: Modelling Mutual Inhibition in textit{Pseudomonas aeruginosa} Sibling Colonies
Pith reviewed 2026-05-15 02:36 UTC · model grok-4.3
The pith
Sibling colonies of Pseudomonas aeruginosa separate because local nutrient depletion slows growth and motility in a self-reinforcing way.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The authors establish that mutual inhibition between sibling colonies results from localized nutrient depletion coupled to a dynamic feedback between growth and motility. This mechanism accounts for the observed sharp demarcation lines and the dependence on initial nutrient levels, without requiring direct inhibition or communication.
What carries the argument
The nutrient-motility feedback, in which nutrient scarcity at colony edges reduces growth rates and modifies motility to limit further advance toward depleted zones.
If this is right
- Colony separation strength decreases as initial nutrient concentration increases.
- The effect persists in isogenic strains without quorum sensing or lethal factors.
- Quantitative density profiles match model predictions for boundary sharpness.
- The framework applies to spatial dynamics in other motile microbial systems.
Where Pith is reading between the lines
- Similar nutrient-based avoidance might operate in natural biofilms or infections where resources are patchy.
- Controlling nutrient gradients could be used to direct bacterial spatial arrangements in synthetic microbiomes.
- Testing the model in gels with varying stiffness would confirm if mechanics play any secondary role.
Load-bearing premise
That ruling out gel compression, lethal inhibition, and quorum sensing leaves nutrient depletion and motility feedback as the sole drivers of the separation.
What would settle it
If colonies still form sharp lines when nutrients are supplied uniformly and in excess, or if disabling motility eliminates the inhibition while keeping growth intact.
Figures











read the original abstract
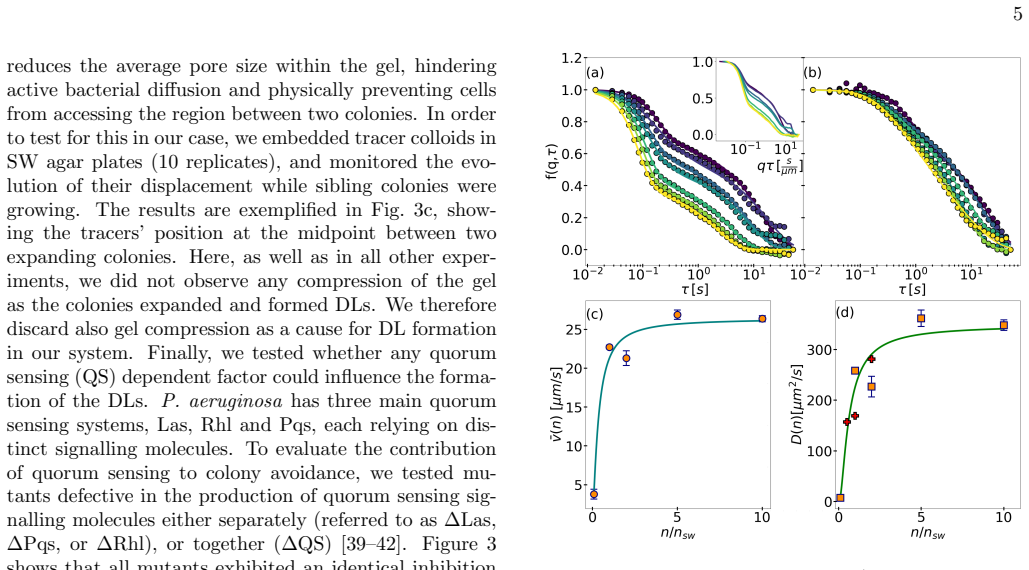
The striking variety of macroscopic morphologies displayed by bacterial colonies depends on microscopic environmental and behavioural details in a manner that is currently not well understood. A surprising example is sibling inhibition, whereby isogenic bacterial colonies spreading in soft agar hydrogels tend to avoid each other and form sharp demarcation lines when growing nearby. Here we investigate this effect with the common pathogen \textit{Pseudomonas aeruginosa}, by combining quantitative density measurements with a minimal biophysical model. Our results show that the phenomenon does not depend on gel compression, lethal inhibition or quorum sensing-dependent cell communication. Instead, colony separation is driven by localised nutrient depletion through a dynamic feedback between growth and motility. The model, which is calibrated using experimental data, captures key observations including the dependence of inhibition strength on the initial nutrient concentration. This work establishes nutrient availability and non-lethal motility inhibition as central factors underlying sibling inhibition, providing a generalisable framework for microbial spatial dynamics with implications for understanding bacterial interactions in tissues, soils and engineered microbiomes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript investigates sibling inhibition in Pseudomonas aeruginosa colonies spreading in soft agar, where isogenic colonies form sharp demarcation lines. Combining quantitative density measurements with a minimal biophysical model, the authors conclude that the effect is independent of gel compression, lethal inhibition, and quorum sensing; instead, colony separation arises from localized nutrient depletion that creates a dynamic feedback between growth and motility. The model is calibrated to experimental density data and reproduces the observed dependence of inhibition strength on initial nutrient concentration.
Significance. If the nutrient-motility feedback mechanism is confirmed, the work supplies a generalizable biophysical framework for microbial spatial self-organization in active matter. It identifies nutrient availability and non-lethal motility inhibition as key drivers of colony morphology, with direct implications for bacterial interactions in tissues, soils, and synthetic microbiomes. The quantitative experimental-modeling approach strengthens the case for nutrient-dependent kinematic exclusion over alternative explanations.
major comments (1)
- [Model calibration and validation sections] The central claim that localized nutrient depletion drives colony separation via growth-motility feedback rests on model agreement with density profiles and exclusion of alternatives, but lacks direct local nutrient measurements in the inter-colony region at the relevant times and length scales. Without observed depletion gradients (or an independent, parameter-free prediction of the effect), other non-lethal density- or motility-dependent mechanisms consistent with the same data cannot be ruled out, weakening the load-bearing inference.
minor comments (1)
- Figure legends and axis labels in the density-profile panels would benefit from explicit indication of error bars and the number of biological replicates to improve quantitative readability.
Simulated Author's Rebuttal
We thank the referee for their constructive review and for highlighting the need for stronger validation of the nutrient-depletion mechanism. We address the major comment below and clarify the evidential basis of our conclusions while remaining faithful to the data and model presented in the manuscript.
read point-by-point responses
-
Referee: [Model calibration and validation sections] The central claim that localized nutrient depletion drives colony separation via growth-motility feedback rests on model agreement with density profiles and exclusion of alternatives, but lacks direct local nutrient measurements in the inter-colony region at the relevant times and length scales. Without observed depletion gradients (or an independent, parameter-free prediction of the effect), other non-lethal density- or motility-dependent mechanisms consistent with the same data cannot be ruled out, weakening the load-bearing inference.
Authors: We acknowledge that direct, spatially resolved nutrient measurements in the inter-colony gap at the relevant length and time scales would constitute stronger, more direct evidence. Such measurements are technically demanding in soft-agar setups and were not performed in this study. Our inference instead rests on three interlocking lines of evidence that are all present in the manuscript: (i) quantitative density profiles measured across multiple initial nutrient concentrations, (ii) explicit experimental exclusion of gel compression, lethality, and quorum-sensing communication, and (iii) a minimal biophysical model whose parameters are fixed by the density data and that quantitatively reproduces the observed dependence of demarcation-line strength on nutrient level. Because the model contains no free parameters once calibrated and still captures the nutrient dependence, we regard the agreement as non-trivial support for the growth-motility feedback. We will add a dedicated paragraph in the revised Discussion that explicitly states this limitation and outlines feasible routes for future direct nutrient imaging. revision: partial
- Direct local nutrient concentration measurements in the inter-colony region at the relevant times and length scales
Circularity Check
Model calibrated to density data reproduces nutrient dependence by construction
specific steps
-
fitted input called prediction
[Abstract]
"The model, which is calibrated using experimental data, captures key observations including the dependence of inhibition strength on the initial nutrient concentration."
Parameters are fitted to quantitative density measurements; the same model is then said to 'capture' the nutrient-concentration dependence. This dependence is part of the calibration data or directly implied by it, so the reported agreement is forced by the fit rather than a genuine out-of-sample prediction of the dynamic feedback.
full rationale
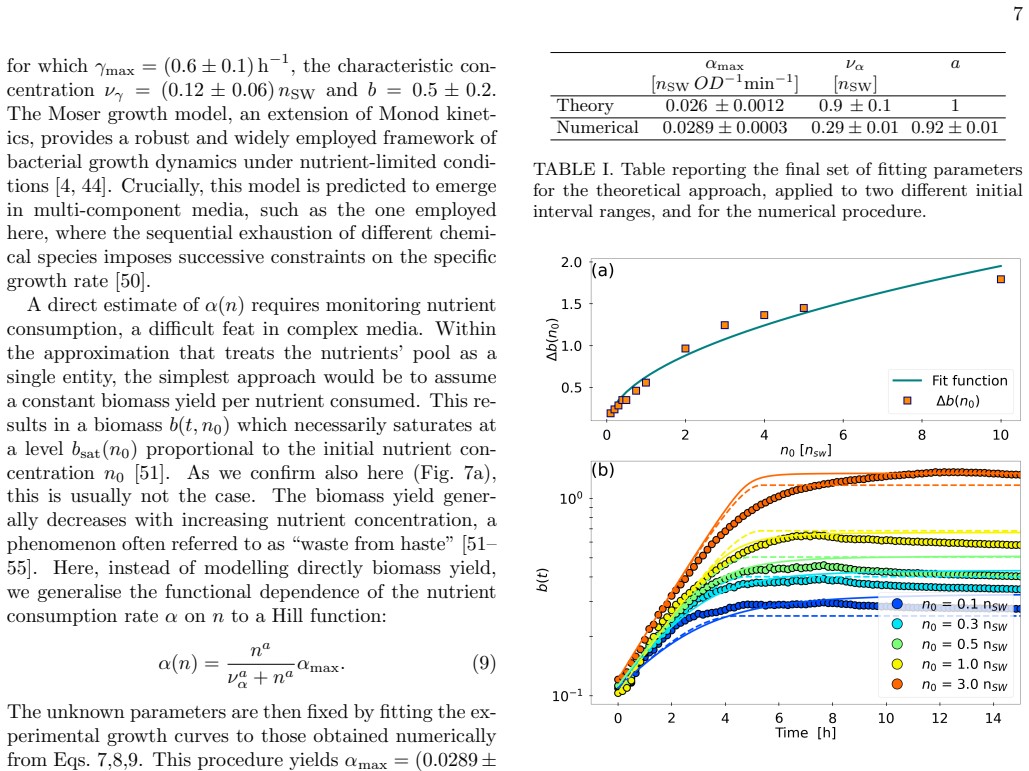
The paper's central claim rests on a minimal biophysical model whose parameters are fitted to experimental density profiles; this same model is then reported to capture the observed dependence of inhibition strength on initial nutrient concentration. Because the reproduction uses the fitted inputs, the agreement is expected rather than an independent test of the nutrient-motility feedback mechanism. No parameter-free predictions or direct local nutrient measurements are shown, so the load-bearing inference remains partially circular. The exclusion of alternative mechanisms (gel compression, quorum sensing) is stated but does not remove the calibration dependence.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Y. M. Bar-On, R. Phillips, and R. Milo, Proceedings of the National Academy of Sciences115, 6506 (2018)
work page 2018
- [2]
- [3]
-
[4]
T. Bhattacharjee, D. B. Amchin, J. A. Ott, F. Kratz, and S. S. Datta, Biophysical Journal120, 3483 (2021)
work page 2021
-
[5]
A. Wolfe and H. Berg, Proceedings of the National Academy of Sciences of the United States of America 86, 6973 (1989)
work page 1989
-
[6]
S. S. Datta, A. P. Steinberg, and R. F. Ismagilov, Pro- ceedings of the National Academy of Sciences113, 7041 (2016). 13
work page 2016
- [7]
-
[8]
B. J. Toley and N. S. Forbes, Integrative Biology4, 165 (2011)
work page 2011
- [9]
- [10]
- [11]
-
[12]
K. A. Gibbs, M. L. Urbanowski, and E. P. Greenberg, Science321, 256 (2008)
work page 2008
-
[13]
P. Stefanic, B. Kraigher, N. A. Lyons, R. Kolter, and I. Mandic-Mulec, Proceedings of the National Academy of Sciences112, 14042 (2015)
work page 2015
-
[14]
N. A. Lyons, B. Kraigher, P. Stefanic, I. Mandic-Mulec, and R. Kolter, Current Biology26, 733 (2016)
work page 2016
-
[15]
A. Mart´ ınez-Calvo, C. Trenado-Yuste, H. Lee, J. Gore, N. S. Wingreen, and S. S. Datta, Phys. Rev. X15, 011016 (2025)
work page 2025
- [16]
- [17]
- [18]
-
[19]
E. L. Munson, M. A. Pfaller, and G. V. Doern, Journal of Clinical Microbiology40, 4285 (2002)
work page 2002
-
[20]
D. R. Espeso, E. Mart´ ınez-Garc´ ıa, V. de Lorenzo, and ´A. Go˜ ni Moreno, Frontiers in Microbiology7, 1437 (2016)
work page 2016
- [21]
-
[22]
A. Sekowska, J.-B. Masson, A. Celani, A. Danchin, and M. Vergassola, Biophysical Journal97, 688 (2009)
work page 2009
-
[23]
R. Paul, T. Ghosh, T. Tang, and A. Kumar, Soft Matter 15, 5400 (2019)
work page 2019
-
[24]
A. Curatolo, N. Zhou, Y. Zhao, A. Daerr, J. Tailleur, and J. Huang, Nature Physics16, 1 (2020)
work page 2020
- [25]
-
[26]
R. Cruz-L´ opez, P. Kolesi´ nski, F. De Boever, D. Green, M. Carrano, and C. Carrano, Microorganisms9, 103 (2021)
work page 2021
-
[27]
M. Cervoni, D. Sposato, A. Lo Sciuto, and F. Imperi, Antibiotics12, 200 (2023)
work page 2023
-
[28]
D. Sposato, J. Mercolino, L. Torrini, P. Sperandeo, M. Lucidi, R. Alegiani, I. Varone, G. Molesini, L. Leoni, G. Rampioni, P. Visca, and F. Imperi, mSphere9, e0067723 (2024)
work page 2024
-
[29]
M. Cervoni, A. Ferriero, A. Lo Sciuto, F. Guidi, N. Babic Jordamovic, S. Piazza, O. Jousson, A. Esposito, and F. Imperi, Antibiotics14, 601 (2025)
work page 2025
-
[30]
M. Lichtenberg, T. H. Jakobsen, M. K¨ uhl, M. Kolpen, P. Ø. Jensen, and T. Bjarnsholt, FEMS Microbiology Re- views46, fuac018 (2022)
work page 2022
- [31]
- [32]
-
[33]
J. Schwarz-Linek, J. Arlt, A. Jepson, A. Dawson, T. Vis- sers, D. Miroli, T. Pilizota, V. A. Martinez, and W. C. Poon, Colloids and Surfaces B: Biointerfaces137, 2 (2016), biocolloids and Colloids in Biology
work page 2016
-
[34]
L. G. Wilson, V. A. Martinez, J. Schwarz-Linek, J. Tailleur, G. Bryant, P. N. Pusey, and W. C. K. Poon, Phys. Rev. Lett.106, 018101 (2011)
work page 2011
-
[35]
D. Germain, M. Leocmach, and T. Gibaud, American Journal of Physics84, 202 (2016)
work page 2016
-
[36]
V. A. Martinez, R. Besseling, O. A. Croze, J. Tailleur, M. Reufer, J. Schwarz-Linek, L. G. Wilson, M. A. Bees, and W. C. K. Poon, Biophysical Journal103, 1637 (2012)
work page 2012
-
[37]
E. Lattuada, F. Krautgasser, M. Lavaud, F. Giavazzi, and R. Cerbino, The Journal of Chemical Physics163, 161501 (2025)
work page 2025
- [38]
-
[39]
G. Rampioni, C. Pustelny, M. P. Fletcher, V. J. Wright, M. Bruce, K. P. Rumbaugh, S. Heeb, M. C´ amara, and P. Williams, Environmental Microbiology12, 1659 (2010)
work page 2010
-
[40]
M. Letizia, M. Mellini, A. Fortuna, P. Visca, F. Imperi, L. Leoni, and G. Rampioni, Microbiology Spectrum10, e00961 (2022)
work page 2022
-
[41]
M. Mellini, M. Letizia, L. Caruso, A. Guiducci, C. Meneghini, S. Heeb, P. Williams, M. C´ amara, P. Visca, F. Imperi, L. Leoni, and G. Rampioni, mBio 14, e02039 (2023)
work page 2023
-
[42]
R. Bond` ı, F. Longo, M. Messina, F. D’Angelo, P. Visca, L. Leoni, and G. Rampioni, Molecular BioSystems (MBS)13, 1080 (2017)
work page 2017
-
[43]
A. W. C. Lau and T. C. Lubensky, Phys. Rev. E76, 011123 (2007)
work page 2007
- [44]
-
[45]
K. Martens, L. Angelani, R. Di Leonardo, and L. Boc- quet, The European physical journal. E, Soft matter35, 9761 (2012)
work page 2012
-
[46]
Monod, Annual Review of Microbiology3, 371 (1949)
J. Monod, Annual Review of Microbiology3, 371 (1949)
work page 1949
-
[47]
Moser, Cold Spring Harbor symposia on quantitative biology22, 121 (1957)
H. Moser, Cold Spring Harbor symposia on quantitative biology22, 121 (1957)
work page 1957
- [48]
-
[49]
J. F. Yamagishi and T. S. Hatakeyama, Proceedings of the National Academy of Sciences122, e2515031122 (2025)
work page 2025
-
[50]
J. Yamagishi and T. Hatakeyama, Proceedings of the Na- tional Academy of Sciences of the United States of Amer- ica122, e2515031122 (2025)
work page 2025
- [51]
-
[52]
T. Pfeiffer, S. Schuster, and S. Bonhoeffer, Science (New York, N.Y.)292, 504 (2001)
work page 2001
- [53]
-
[54]
J. A. Bonachela, M. Raghib, and S. A. Levin, Proceedings of the National Academy of Sciences108, 20633 (2011)
work page 2011
-
[55]
O. A. Nev, R. J. Lindsay, A. Jepson, L. Butt, R. E. Beardmore, and I. Gudelj, PLOS Computational Biology 14 17, 1 (2021)
work page 2021
-
[56]
C. W. Gardiner,Handbook of Stochastic Methods for Physics, Chemistry and the Natural Sciences, 4th ed., Springer Series in Synergetics, Vol. 13 (Springer, Berlin, Heidelberg, 2009)
work page 2009
-
[57]
P. F. Tupper and X. Yang, Proceedings of the Royal Soci- ety A: Mathematical, Physical and Engineering Sciences 468, 3864 (2012)
work page 2012
-
[58]
J. Arlt, V. Martinez, A. Dawson, T. Pilizota, and W. Poon, Nature Communications9(2018)
work page 2018
-
[59]
G. Frangipane, D. Dell’Arciprete, S. Petracchini, C. Maggi, F. Saglimbeni, S. Bianchi, G. Vizsnyiczai, M. L. Bernardini, and R. Di Leonardo, eLife7, e36608 (2018)
work page 2018
-
[60]
N. Pellicciotta, M. Paoluzzi, D. Buonomo, G. Frangipane, L. Angelani, and R. Leonardo, Nature Communications 14(2023)
work page 2023
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.