Elastohydrodynamic coupling enhances flow generation by coordinated ciliary beating
Pith reviewed 2026-06-28 17:57 UTC · model grok-4.3
The pith
Elastic restoring forces couple with position shifts in cilia to enhance fluid transport, with optimal coordination depending on beat geometry.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Reinforcement learning on the bead-spring cilia model identifies antiplectic coordination as the flow-maximizing state in linear arrays, with the phase difference between neighbours accounting for most enhancement. The analytically tractable tilted-slider model of the weak-coupling limit shows that a shift of the time-averaged position opposite to the effective-stroke direction enhances fluid transport through its coupling with the elastic restoring force. The same model reveals that antiplectic coordination can be optimal, consistent with prior work, whereas symplectic coordination can instead become optimal depending on beat geometry.
What carries the argument
The tilted-slider model, an analytically tractable reduction of the bead-spring cilia dynamics in the weak-coupling limit that tracks how time-averaged position shifts interact with elastic restoring forces to set net flow.
If this is right
- Antiplectic metachronal waves maximize flow in linear arrays for standard beat geometries.
- Phase difference between neighbouring cilia drives most of the transport gain.
- Symplectic coordination can maximize flow when beat geometry is altered in specific ways.
- The elastohydrodynamic mechanism operates through elastic force coupling to average position.
Where Pith is reading between the lines
- The same position-shift mechanism might be tested in two-dimensional ciliary carpets to see whether optimal waves rotate or change character.
- Artificial microfluidic devices could tune beat geometry to switch between antiplectic and symplectic optima without retraining controllers.
- If real biological cilia exhibit symplectic waves in some geometries, the model predicts a measurable shift in their average stroke position.
Load-bearing premise
The weak-coupling limit captured by the tilted-slider model remains representative of the full bead-spring dynamics discovered by reinforcement learning.
What would settle it
A direct numerical simulation or experiment on the bead-spring model with altered beat geometry that produces symplectic coordination as the flow maximum while the tilted-slider model predicts antiplectic would falsify the reduction step.
Figures

read the original abstract
Ciliary arrays pump fluid at low Reynolds number through non-reciprocal beating and phase coordination between neighbouring cilia. Previous studies have demonstrated that antiplectic metachronal waves are more effective than symplectic waves in enhancing transport, and have proposed several physically intuitive explanations for this preference. What remains incomplete is a predictive analytical understanding of how hydrodynamic coupling and beat geometry determine the flow-maximising phase difference. Here, we address this problem in two steps: we first use reinforcement learning to identify flow-maximising coordination in a bead--spring cilia model, and then introduce an analytically tractable reduced model, termed a tilted-slider model, to analyse the weak-coupling limit. Reinforcement learning identifies antiplectic coordination as the flow-maximising state in linear arrays, and shows that the phase difference between neighbouring cilia accounts for most of the flow enhancement. We then use the tilted-slider model to show that a shift of the time-averaged position opposite to the effective-stroke direction enhances fluid transport through its coupling with the elastic restoring force. The reduced model further reveals that antiplectic coordination can be optimal, consistent with previous studies, whereas symplectic coordination can instead become optimal depending on beat geometry. These results identify a simple elastohydrodynamic mechanism underlying flow-maximising metachronal coordination.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that reinforcement learning on a bead-spring cilia model identifies antiplectic metachronal coordination as flow-maximizing in linear arrays, with phase difference between neighbors accounting for most enhancement. A tilted-slider reduced model in the weak-coupling limit is introduced to derive that a time-averaged position shift opposite the effective stroke enhances transport via coupling to the elastic restoring force; the reduced model also shows antiplectic coordination is typically optimal while symplectic can become optimal depending on beat geometry.
Significance. If the reduced model is representative of the full dynamics, the work supplies a predictive analytical mechanism for how elastohydrodynamic coupling and beat geometry set the optimal phase difference, extending prior observations on antiplectic preference. The use of RL for identifying optima and a reduced model for mechanism is a methodological strength.
major comments (2)
- [Tilted-slider model and RL results sections] The central claim requires that the tilted-slider weak-coupling predictions are representative of the bead-spring RL results. No direct quantitative comparison is reported of optimal phase difference, time-averaged displacement, or flow enhancement between the reduced model and the full simulations for matching beat geometry and coupling strength (see the sections presenting RL results and the tilted-slider analysis).
- [Abstract and methods/results on RL] The abstract states that the RL identifies antiplectic coordination and the reduced model derives the position-shift mechanism, but the manuscript provides no equations, error analysis, or validation of the weak-coupling approximation against the full bead-spring dynamics.
Simulated Author's Rebuttal
We thank the referee for their constructive review and positive assessment of the work's significance. We address each major comment below and outline revisions to strengthen the presentation of the reduced model and its connection to the RL results.
read point-by-point responses
-
Referee: [Tilted-slider model and RL results sections] The central claim requires that the tilted-slider weak-coupling predictions are representative of the bead-spring RL results. No direct quantitative comparison is reported of optimal phase difference, time-averaged displacement, or flow enhancement between the reduced model and the full simulations for matching beat geometry and coupling strength (see the sections presenting RL results and the tilted-slider analysis).
Authors: We agree that a direct quantitative comparison would strengthen the central claim. In the revised manuscript we will add a dedicated comparison (new figure and text) for matching beat geometry and weak-coupling strength, reporting the optimal phase difference, time-averaged displacement, and resulting flow enhancement obtained from both the RL bead-spring simulations and the tilted-slider predictions. revision: yes
-
Referee: [Abstract and methods/results on RL] The abstract states that the RL identifies antiplectic coordination and the reduced model derives the position-shift mechanism, but the manuscript provides no equations, error analysis, or validation of the weak-coupling approximation against the full bead-spring dynamics.
Authors: The tilted-slider derivation appears in the methods, yet we acknowledge that the main text lacks explicit governing equations, an error analysis of the approximation, and direct validation. We will move the key equations into the results section, add an error analysis of the weak-coupling limit, and include a validation subsection that compares the reduced-model predictions against the full bead-spring dynamics for corresponding parameters. revision: yes
Circularity Check
No significant circularity; RL discovery and reduced-model analysis remain independent
full rationale
The derivation proceeds in two distinct stages: reinforcement learning supplies an independent numerical identification of flow-maximising states in the bead-spring model, after which the tilted-slider reduced model is introduced separately to analyse the weak-coupling limit and identify the elastohydrodynamic mechanism (time-averaged position shift coupled to elastic force). No equation or claim reduces a 'prediction' to a fitted parameter by construction, no self-citation chain bears the central result, and the optimality statements are not forced by the same quantities used to generate the input states. The chain is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Weak-coupling limit applies to the tilted-slider model
Reference graph
Works this paper leans on
-
[1]
Reinforcement learning In this work, we introduced a reinforcement learning algorithm, proximal policy optimisation (PPO) [47], to search for a strategy to maximise the pumping. Reinforcement learning aims to learn a policy that selects an actionA t based on the current stateS t to maximise the expected cumulative rewardR ep, defined by the immediate rewa...
-
[2]
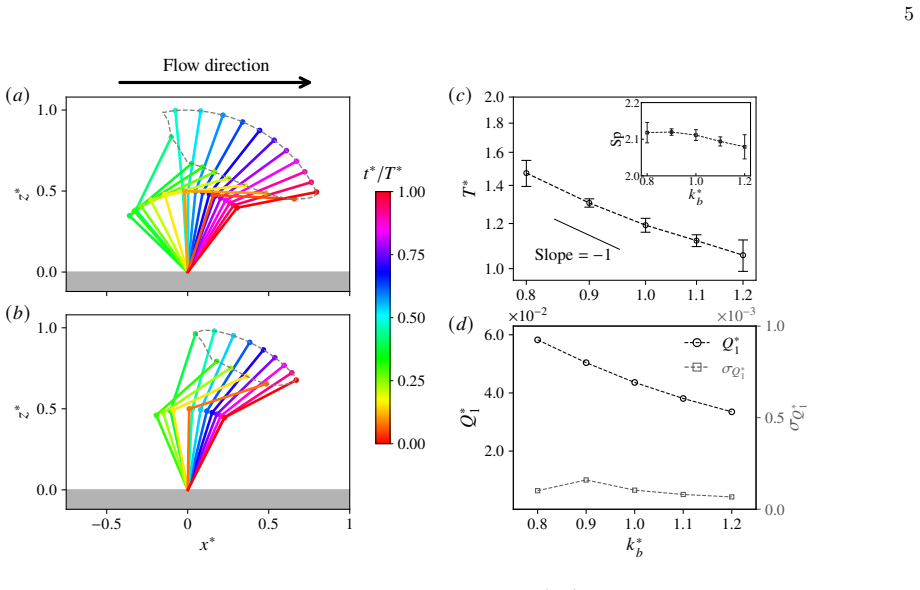
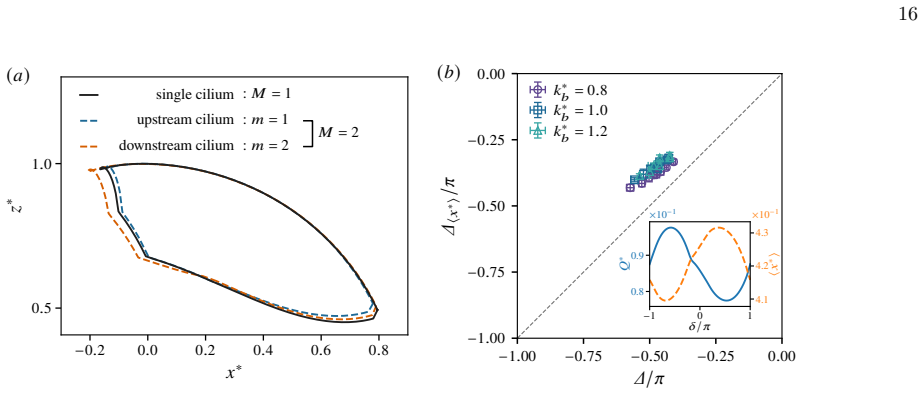
Symbols and error bars in (b,c) show the mean and standard deviation over five independent training runs
for differentM, where Q∗ M andQ ∗ 1 denote the flow rates of the optimisedMcilia and single-cilium systems, respectively. Symbols and error bars in (b,c) show the mean and standard deviation over five independent training runs. The emergence of the optimal beating periodT ∗ can be understood as a consequence of the competition between the elastic relaxati...
-
[3]
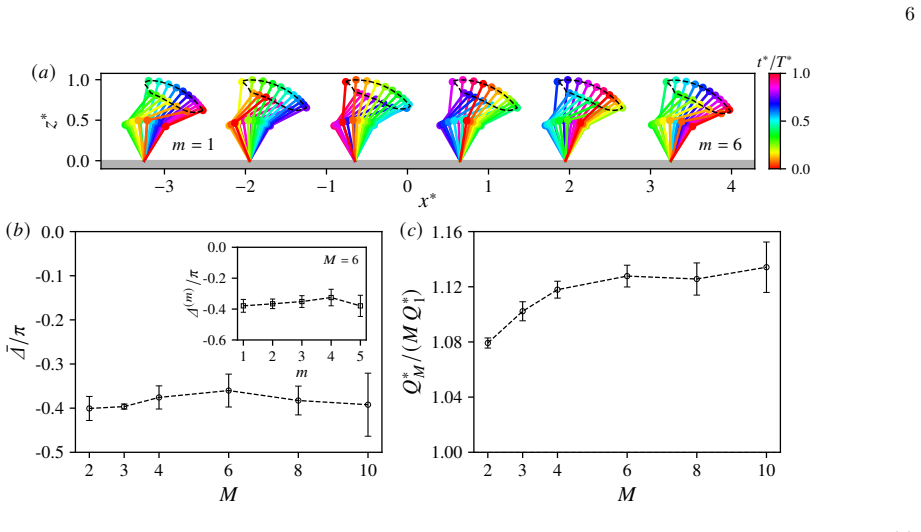
varies with cilia number in figure 3(c), whereQ ∗ 1 is the flow rate of the single cilium obtained in the previous subsection; the quantity Q∗ M /(M Q∗
-
[4]
The figure shows that there is already flow enhancement of 8% even for two cilia, and this ratio increases with the cilia number but saturates at approximately 12% forM≈6
can also be interpreted as the relative flow enhancement due to the coordination. The figure shows that there is already flow enhancement of 8% even for two cilia, and this ratio increases with the cilia number but saturates at approximately 12% forM≈6. Gaugeret al.[24] likewise reported that the flow enhancement ratio saturates, reaching approximately 40...
-
[5]
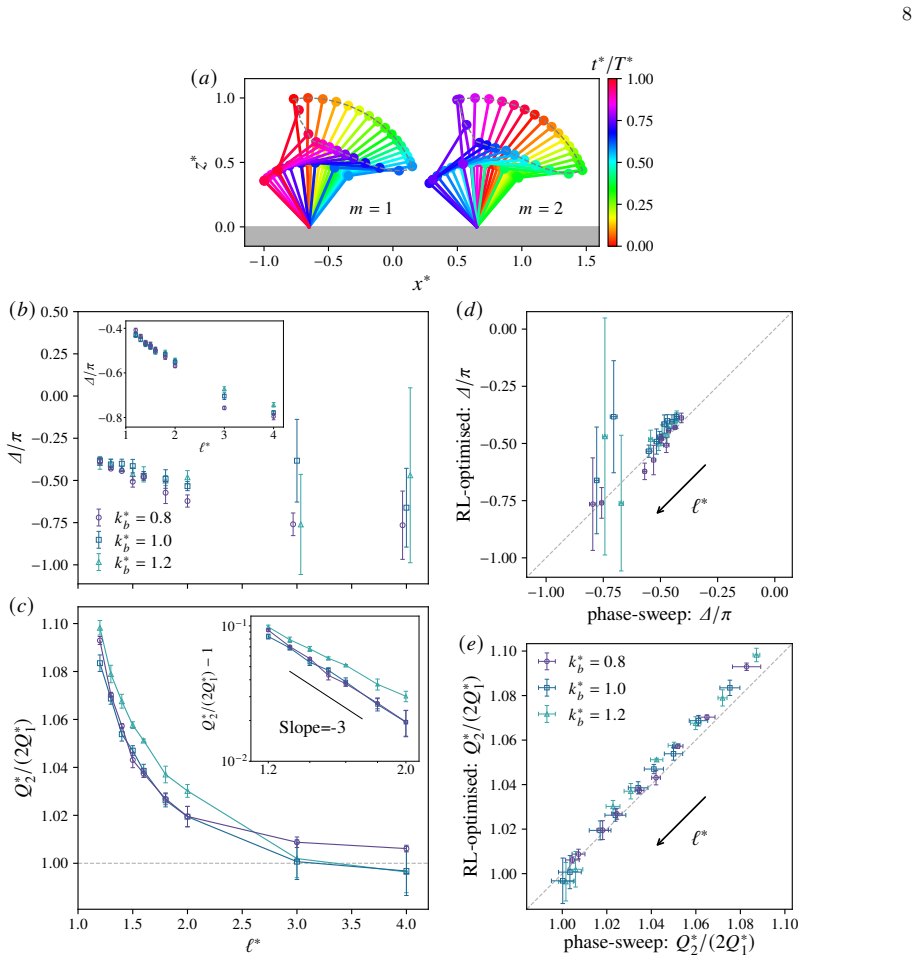
The excess flow ratioQ ∗ 2/(2Q∗ 1)−1 decays rapidly with∼ℓ −3 as shown in the inset
is approximately 10% when the spacing is small and decreases as the spacing increases; for large spacing asℓ ∗ ≳3, the generated flow is well approximated as being proportional to the number of ciliaM. The excess flow ratioQ ∗ 2/(2Q∗ 1)−1 decays rapidly with∼ℓ −3 as shown in the inset. It is interesting to note that both ∆ andQ ∗ 2/(2Q∗
-
[6]
depend only weakly on the bending stiffnessk ∗ b . The results for two cilia naturally raise a more specific question: does the flow enhancement due to the coordination require independent optimisation for each cilium, or can it already be realised by identical beating patterns with an appropriate phase difference? To clarify this point, we perform a simp...
-
[7]
Although reinforcement learning gives slightly largerQ ∗ 2/(2Q∗ 1), this difference remains small, indicating that independent optimisation for each cilium has only a small effect
as reinforcement learning, suggesting that the flow enhancement can be explained primarily by the phase difference rather than by the independent optimisation for each cilium. Although reinforcement learning gives slightly largerQ ∗ 2/(2Q∗ 1), this difference remains small, indicating that independent optimisation for each cilium has only a small effect. ...
-
[8]
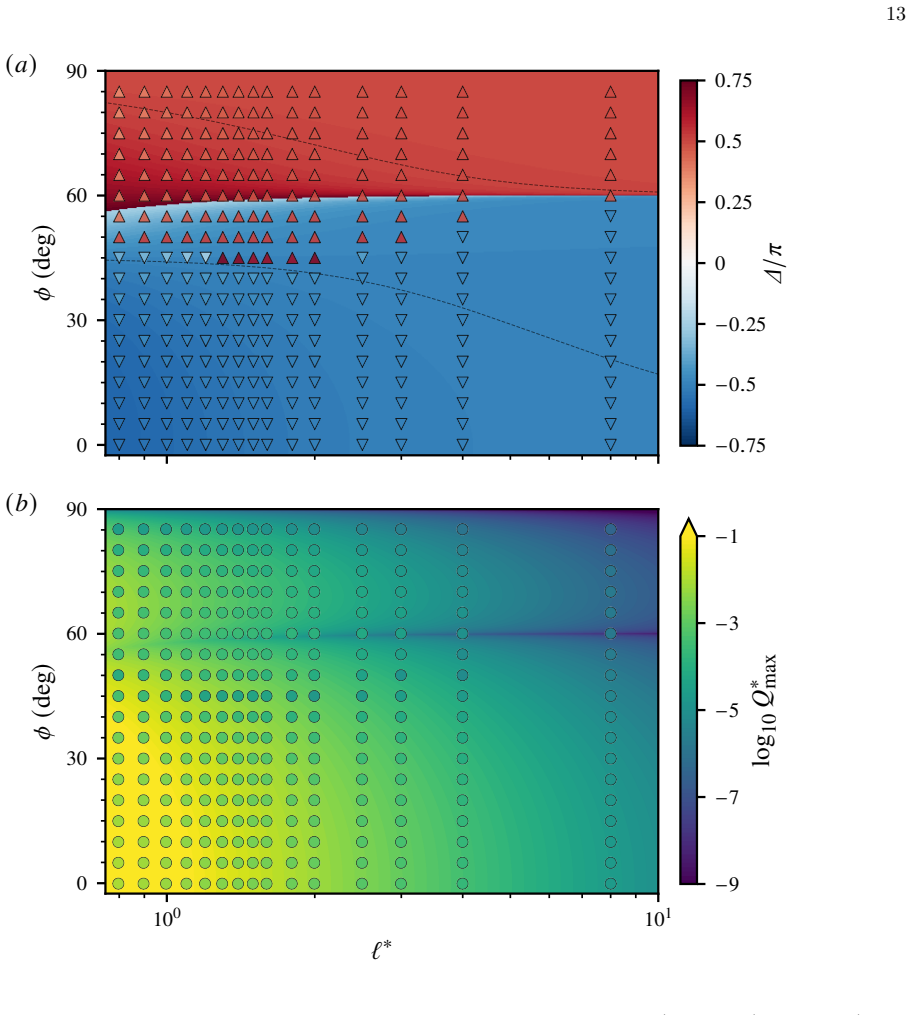
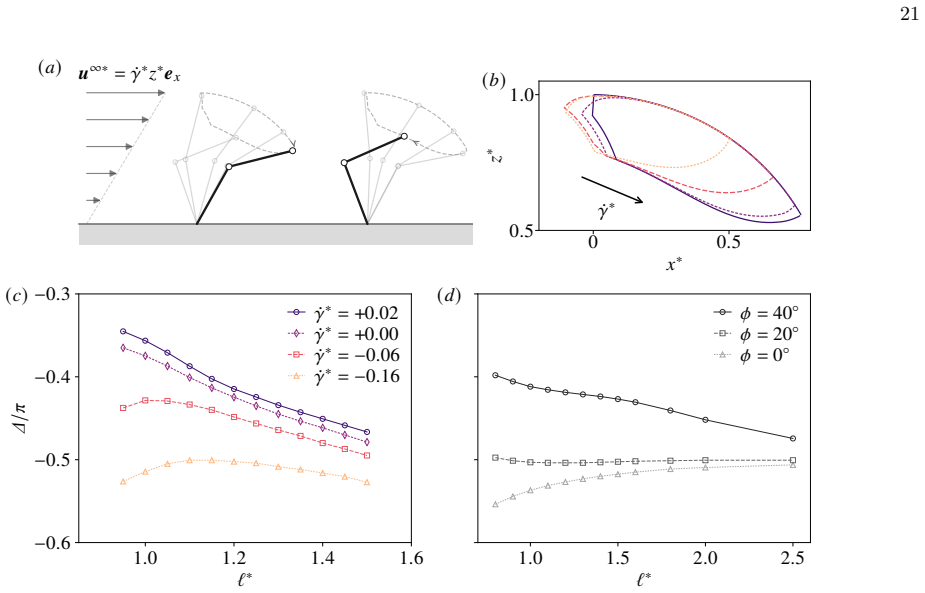
□ 1 Slope=-3 1 2 3 4 /uni2113∗ □0.8 □0.6 □0.4 /u1D6E5//u1D70B (/u1D44E) (/u1D44F) (/u1D450) (/u1D451) (/u1D452) FIG. 4. Beating of two cilia optimised by reinforcement learning. (a) Representative beating patterns fork ∗ b = 0.8 andℓ ∗ = 1.3. (b) Optimal phase difference ∆ as a function of inter-cilium spacingℓ ∗. Inset: the same plot obtained using the p...
-
[9]
Inset: log–log plot of the excess flow ratioQ ∗ 2/(2Q∗ 1)−1
as a function ofℓ ∗. Inset: log–log plot of the excess flow ratioQ ∗ 2/(2Q∗ 1)−1. (d,e) Comparison between RL and the phase-sweep analysis for (d) the flow-rate ratioQ ∗ 2/(2Q∗
-
[10]
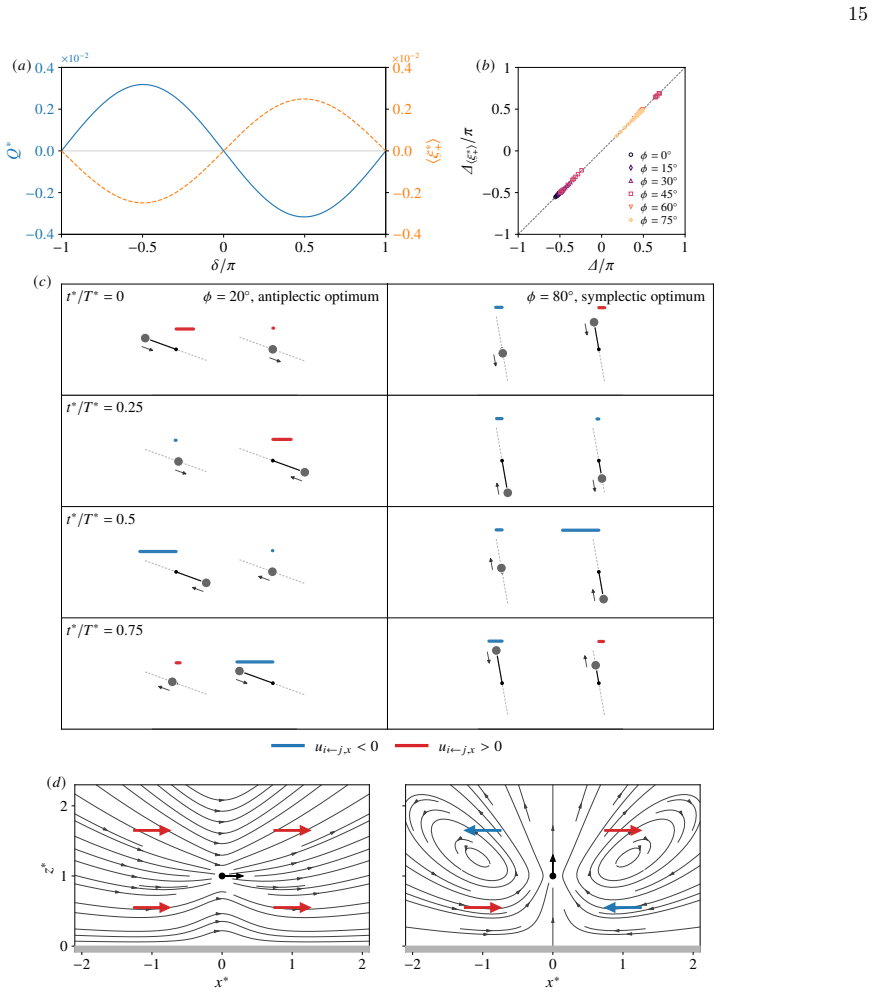
and (e) the optimal imposed offset ∆/π. In (d,e), the vertical error bars indicate the standard deviation over five runs, whereas the horizontal error bars indicate the standard deviation over phase-sweeps using five different torque sequences optimised for a single cilium. IV. MECHANISM OF FLOW ENHANCEMENT In the previous section, reinforcement learning ...
-
[11]
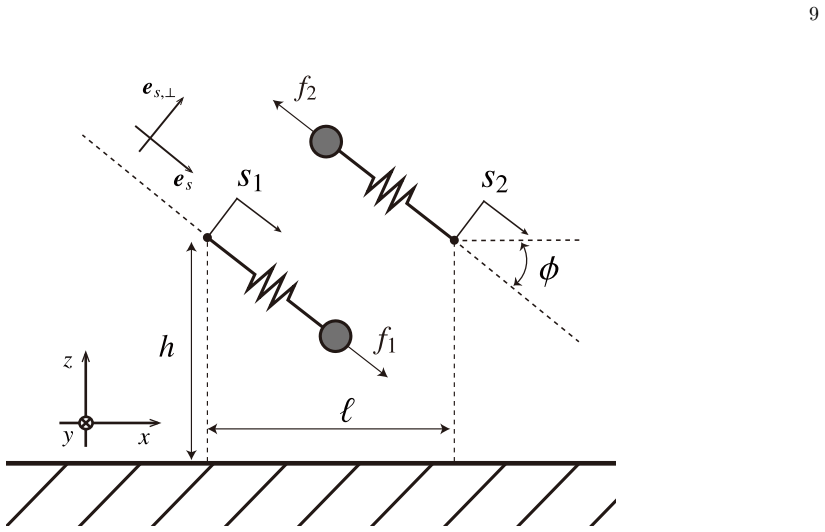
Weak-coupling reduction To simplify the projected dynamics, we introduce the horizontal projections of the slider displacements,ξ∗ i :=s ∗ i cosϕ (i= 1,2), together with the sum and difference variablesξ ∗ + :=ξ ∗ 1 +ξ ∗ 2 andξ ∗ − :=ξ ∗ 2 −ξ ∗
-
[12]
Here,ξ ∗ +/2 is the horizontal shift of the centroid, whileξ ∗ − is the instantaneous difference of the displacement. The bead positions are then written as x∗ 1 =− ℓ∗ 2 +ξ ∗ 1 , z ∗ 1 = 1−ξ ∗ 1 tanϕ, x ∗ 2 = ℓ∗ 2 +ξ ∗ 2 , z ∗ 2 = 1−ξ ∗ 2 tanϕ.(17) Using the far-field expansion of the pair mobility C∗(t∗) := 3 2π z∗ 1 z∗ 2 " cos2 ϕ (ℓ∗ +ξ ∗ −)3 + sin2 ϕ ξ...
-
[13]
Flow-rate expression Using (7), the instantaneous flow rate generated by the two sliders is q∗(t∗) = 1 π 2X i=1 z∗ i f ∗ i cosϕ−k ∗ r ξ∗ i ,(23) and the cycle-averaged dimensionless flow rate is therefore Q∗ = 1 T ∗ Z T ∗ 0 q∗(t∗) dt∗.(24) Substitutingz ∗ i = 1−ξ ∗ i tanϕand rewriting the result in terms ofξ ∗ ± andf ∗ ±, we obtain Q∗ = 1 πT ∗ Z T ∗ 0 −k∗...
-
[14]
Zeroth-order response SinceC ∗ ∼O(ε), substituting the regular expansion (22) into (19) and collecting theO(1) terms yields ˙ξ∗ ±,0 +γ ∗ 0 k∗ r ξ∗ ±,0 =γ ∗ 0 f ∗ ± cosϕ.(30) Thus, at leading-order, the two sliders are decoupled and each mode behaves as a forced first-order relaxation process. From (30), the forcing terms are f ∗ +(t∗) = 2 cos δ 2 cos ω∗t∗...
-
[15]
Since theO(ε) correction to (29) depends on the mean shift ofξ ∗ +, it is sufficient here to consider the equation forξ ∗ +,1
First-order phase-dependent flow rate We now turn to the first non-trivial contribution to the cycle-averaged transport. Since theO(ε) correction to (29) depends on the mean shift ofξ ∗ +, it is sufficient here to consider the equation forξ ∗ +,1. Retaining termsO(ε) in (19) gives ˙ξ∗ +,1 +γ ∗ 0 k∗ r ξ∗ +,1 =C ∗ f ∗ + cosϕ−k ∗ r ξ∗ +,0 = C∗ γ∗ 0 ˙ξ∗ +,0,(...
-
[16]
Expanding in powers ofz ∗/r∗ ∥ andz ∗ 0 /r∗ ∥, one obtains G∗ 11(r∗,r ∗
Far-field expansion of the Blake-tensor From the observation pointr= (x,0, z) and source pointr 0 = (x0,0, z 0), we define ˆx:=x−x 0, r ∥ :=|ˆx|.(B1) In the far-field regime, max{z ∗, z∗ 0 } ≪r ∗ ∥, the free-space Stokeslet and its image in the Blake representation cancel at O((r∗ ∥)−1), so that the leading wall-mediated contribution arises only at higher...
-
[17]
= 3 2π z∗z∗ 0 (ˆx∗)2 (r∗ ∥)5 +O((r ∗ ∥)−5),(B2) G∗ 13(r∗,r ∗
-
[18]
=− 3 2π z∗(z∗ 0)2 ˆx∗ (r∗ ∥)5 +O((r ∗ ∥)−6),(B3) G∗ 31(r∗,r ∗
-
[19]
= 3 2π (z∗)2z∗ 0 ˆx∗ (r∗ ∥)5 +O((r ∗ ∥)−6),(B4) while G∗ 33(r∗,r ∗
-
[20]
=O((r ∗ ∥)−5).(B5) Note that they-components ofGwere omitted here because they vanish and therefore do not contribute to the dynamics. The scaling of these terms will be important below in obtaining the asymptotic expression of the pair mobility;G ∗ 11 has the lowest leading-orderO((r ∗ ∥)−3), followed byG ∗ 13 andG ∗ 31 withO((r ∗ ∥)−4), andG ∗ 33 appear...
-
[21]
Pair mobility in the tilted-slider geometry The tangential force acting on thej-th bead is F ∗,∥ j = f ∗ j −k ∗ r s∗ j es (B6) wheree s = (cosϕ,0,−sinϕ) is the slider axis. Note that the normal constraint force Λ, which appears in equation (16), is neglected since Λ =O((ℓ ∗)−3) would contribute only anO((ℓ ∗)−6) correction to the pair mobility, once we as...
-
[22]
={G ∗(r∗ 1,r ∗ 2)}T, and henceeC∗(r∗ 2,r ∗
-
[23]
Using the slider geometry (17), one has ˆx∗ =x ∗ 1 −x ∗ 2 =−(ℓ ∗ +ξ ∗ −), r ∗ ∥ =ℓ ∗ +ξ ∗ − >0, z ∗ 2 −z ∗ 1 =−ξ ∗ − tanϕ.(B11) Substituting (B11) into (B2) gives G∗ 11(r∗ 1,r ∗
= eC∗(r∗ 1,r ∗ 2). Using the slider geometry (17), one has ˆx∗ =x ∗ 1 −x ∗ 2 =−(ℓ ∗ +ξ ∗ −), r ∗ ∥ =ℓ ∗ +ξ ∗ − >0, z ∗ 2 −z ∗ 1 =−ξ ∗ − tanϕ.(B11) Substituting (B11) into (B2) gives G∗ 11(r∗ 1,r ∗
-
[24]
= 3 2π z∗ 1 z∗ 2 (ℓ∗ +ξ ∗ −)3 +O((r ∗ ∥)−5),(B12) while combining (B3) and (B4) yields G∗ 13(r∗ 1,r ∗
-
[25]
= 3 2π z∗ 1 z∗ 2(z∗ 2 −z ∗ 1) (ℓ∗ +ξ ∗ −)4 +O((r ∗ ∥)−6).(B13) Substituting (B12) and (B13) into (B9), and omitting theG ∗ 33 contribution and the higher-order far-field remainders, the projected pair mobility can be written, at the order retained here, as eC∗(r∗ 1,r ∗
-
[26]
The calculation is straight- forward but lengthy, and is therefore collected here
=C ∗(t∗) +O((r ∗ ∥)−5), where C∗(t∗) := 3 2π z∗ 1 z∗ 2 " cos2 ϕ (ℓ∗ +ξ ∗ −)3 + sin2 ϕ ξ∗ − (ℓ∗ +ξ ∗ −)4 # .(B14) Appendix C: Evaluation of the first-order mean flux In this appendix, we summarize the analytical evaluation of the period average in (39). The calculation is straight- forward but lengthy, and is therefore collected here. We begin from the lea...
-
[27]
=c 3 cosϑ ∗ +c 4 sin 2ϑ∗ +c 5 cos 3ϑ∗, 1 ω∗ ˙W ∗ 0 =c 6 cosϑ ∗ +c 7 sin 2ϑ∗, (C8) where c0 = 1 + tan2 ϕ 8 A∗2 c −A ∗2 s , c 1 = tanϕ A ∗ c , c 2 =− tan2 ϕ 8 A∗2, c3 =−tanϕ A ∗ s 1 + tan2 ϕ 16 A∗2 c −3A ∗2 s , c 4 =− tan2 ϕ 2 A∗ c A∗ s, c 5 = tan3 ϕ 16 A∗2A∗ s, c6 =−A ∗ c , c 7 =− tanϕ 4 A∗2. (C9) Reducing the remaining trigonometric polynomials to harmoni...
-
[28]
J. R. Blake and M. A. Sleigh, Mechanics of ciliary locomotion, Biological Reviews49, 85 (1974)
1974
-
[29]
Lauga and T
E. Lauga and T. R. Powers, The hydrodynamics of swimming microorganisms, Reports on progress in physics72, 096601 (2009)
2009
-
[30]
Gilpin, M
W. Gilpin, M. S. Bull, and M. Prakash, The multiscale physics of cilia and flagella, Nature Reviews Physics2, 74 (2020)
2020
-
[31]
Omori and T
T. Omori and T. Ishikawa, Ciliary fluid dynamics of swimming, feeding, pumping, and sensing, Physical Review Fluids 10, 080501 (2025)
2025
-
[32]
Purcell, Life at low reynolds number, American Journal of Physics45, 3 (1977)
E. Purcell, Life at low reynolds number, American Journal of Physics45, 3 (1977)
1977
-
[33]
Ishimoto and M
K. Ishimoto and M. Yamada, A coordinate-based proof of the scallop theorem, SIAM Journal on Applied Mathematics 72, 1686 (2012)
2012
-
[34]
Aiello and M
E. Aiello and M. A. Sleigh, The metachronal wave of lateral cilia of mytilus edulis, The Journal of cell biology54, 493 (1972)
1972
-
[35]
M. J. Sanderson and M. A. Sleigh, Ciliary activity of cultured rabbit tracheal epithelium: beat pattern and metachrony, Journal of cell science47, 331 (1981)
1981
-
[36]
Machemer, Ciliary activity and the origin of metachrony in paramecium: effects of increased viscosity, Journal of Experimental Biology57, 239 (1972)
H. Machemer, Ciliary activity and the origin of metachrony in paramecium: effects of increased viscosity, Journal of Experimental Biology57, 239 (1972)
1972
-
[37]
Gheber and Z
L. Gheber and Z. Priel, Synchronization between beating cilia, Biophysical journal55, 183 (1989)
1989
-
[38]
D. R. Brumley, M. Polin, T. J. Pedley, and R. E. Goldstein, Hydrodynamic synchronization and metachronal waves on the surface of the colonial alga volvox carteri, Phys. Rev. Lett.109, 268102 (2012)
2012
-
[39]
Knight-Jones, Relations between metachronism and the direction of ciliary beat in metazoa, Journal of Cell Science3, 503 (1954)
E. Knight-Jones, Relations between metachronism and the direction of ciliary beat in metazoa, Journal of Cell Science3, 503 (1954)
1954
-
[40]
M. C. Lagomarsino, B. Bassetti, and P. Jona, Rowers coupled hydrodynamically. modeling possible mechanisms for the cooperation of cilia, The European Physical Journal B-Condensed Matter and Complex Systems26, 81 (2002)
2002
-
[41]
Wollin and H
C. Wollin and H. Stark, Metachronal waves in a chain of rowers with hydrodynamic interactions, The European Physical Journal E34, 42 (2011)
2011
-
[42]
Vilfan and F
A. Vilfan and F. J¨ ulicher, Hydrodynamic flow patterns and synchronization of beating cilia, Physical review letters96, 058102 (2006)
2006
-
[43]
Niedermayer, B
T. Niedermayer, B. Eckhardt, and P. Lenz, Synchronization, phase locking, and metachronal wave formation in ciliary chains, Chaos: An Interdisciplinary Journal of Nonlinear Science18(2008)
2008
-
[44]
Uchida and R
N. Uchida and R. Golestanian, Generic conditions for hydrodynamic synchronization, Physical Review Letters106, 058104 (2011)
2011
-
[45]
Uchida and R
N. Uchida and R. Golestanian, Synchronization and collective dynamics in a carpet of microfluidic rotors, Physical review letters104, 178103 (2010)
2010
-
[46]
F. Meng, R. R. Bennett, N. Uchida, and R. Golestanian, Conditions for metachronal coordination in arrays of model cilia, Proceedings of the National Academy of Sciences118, e2102828118 (2021)
2021
-
[47]
A. V. Kanale, F. Ling, H. Guo, S. F¨ urthauer, and E. Kanso, Spontaneous phase coordination and fluid pumping in model ciliary carpets, Proceedings of the National Academy of Sciences119, e2214413119 (2022)
2022
-
[48]
von Kenne, M
A. von Kenne, M. B¨ ar, and T. Niedermayer, Hydrodynamic synchronization of elastic cilia: How surface effects determine the characteristics of metachronal waves, Physical Review E109, 054407 (2024)
2024
-
[49]
Cheng, A
Z. Cheng, A. Vilfan, Y. Wang, R. Golestanian, and F. Meng, Near-field hydrodynamic interactions determine travelling wave directions of collectively beating cilia, Journal of the Royal Society Interface21(2024)
2024
-
[50]
Chakrabarti, S
B. Chakrabarti, S. F¨ urthauer, and M. J. Shelley, A multiscale biophysical model gives quantized metachronal waves in a lattice of beating cilia, Proceedings of the National Academy of Sciences119, e2113539119 (2022)
2022
-
[51]
E. M. Gauger, M. T. Downton, and H. Stark, Fluid transport at low reynolds number with magnetically actuated artificial cilia, The European Physical Journal E28, 231 (2009)
2009
-
[52]
Khaderi, J
S. Khaderi, J. Den Toonder, and P. Onck, Microfluidic propulsion by the metachronal beating of magnetic artificial cilia: a numerical analysis, Journal of fluid mechanics688, 44 (2011). 23
2011
-
[53]
Y. Ding, J. C. Nawroth, M. J. McFall-Ngai, and E. Kanso, Mixing and transport by ciliary carpets: a numerical study, Journal of Fluid Mechanics743, 124 (2014)
2014
-
[54]
Zhang, J
R. Zhang, J. den Toonder, and P. R. Onck, Transport and mixing by metachronal waves in nonreciprocal soft robotic pneumatic artificial cilia at low reynolds numbers, Physics of Fluids33(2021)
2021
-
[55]
Zhang, J
R. Zhang, J. den Toonder, and P. R. Onck, Metachronal patterns by magnetically-programmable artificial cilia surfaces for low reynolds number fluid transport and mixing, Soft Matter18, 3902 (2022)
2022
-
[56]
Milana, R
E. Milana, R. Zhang, M. R. Vetrano, S. Peerlinck, M. De Volder, P. R. Onck, D. Reynaerts, and B. Gorissen, Metachronal patterns in artificial cilia for low reynolds number fluid propulsion, Science advances6, eabd2508 (2020)
2020
-
[57]
X. Dong, G. Z. Lum, W. Hu, R. Zhang, Z. Ren, P. R. Onck, and M. Sitti, Bioinspired cilia arrays with programmable nonreciprocal motion and metachronal coordination, Science advances6, eabc9323 (2020)
2020
-
[58]
Chateau, J
S. Chateau, J. Favier, U. D’ortona, and S. Poncet, Transport efficiency of metachronal waves in 3d cilium arrays immersed in a two-phase flow, Journal of Fluid Mechanics824, 931 (2017)
2017
-
[59]
Chateau, U
S. Chateau, U. d’Ortona, S. Poncet, and J. Favier, Transport and mixing induced by beating cilia in human airways, Frontiers in physiology9, 161 (2018)
2018
-
[60]
Chateau, J
S. Chateau, J. Favier, S. Poncet, and U. d’Ortona, Why antiplectic metachronal cilia waves are optimal to transport bronchial mucus, Physical Review E100, 042405 (2019)
2019
-
[61]
H. Ito, T. Omori, and T. Ishikawa, Swimming mediated by ciliary beating: comparison with a squirmer model, Journal of Fluid Mechanics874, 774 (2019)
2019
-
[62]
Omori, H
T. Omori, H. Ito, and T. Ishikawa, Swimming microorganisms acquire optimal efficiency with multiple cilia, Proceedings of the National Academy of Sciences117, 30201 (2020)
2020
-
[63]
Z. Zou, Y. Liu, Y.-N. Young, O. S. Pak, and A. C. Tsang, Gait switching and targeted navigation of microswimmers via deep reinforcement learning, Communications Physics5, 158 (2022)
2022
-
[64]
L.-S. Lin, K. Yasuda, K. Ishimoto, and S. Komura, Emergence of odd elasticity in a microswimmer using deep reinforcement learning, Physical Review Research6, 033016 (2024)
2024
-
[65]
Tokoro, H
F. Tokoro, H. Takayama, S. Deguchi, A. Z¨ ottl, and D. Matsunaga, Optimal undulatory swimming with constrained deformation and actuation intervals, Physical Review Fluids11, 023102 (2026)
2026
-
[66]
Hamilton and P
E. Hamilton and P. Cicuta, Changes in geometrical aspects of a simple model of cilia synchronization control the dynamical state, a possible mechanism for switching of swimming gaits in microswimmers, PloS one16, e0249060 (2021)
2021
-
[67]
Y. W. Kim and R. R. Netz, Pumping fluids with periodically beating grafted elastic filaments, Physical review letters96, 158101 (2006)
2006
-
[68]
Osterman and A
N. Osterman and A. Vilfan, Finding the ciliary beating pattern with optimal efficiency, Proceedings of the National Academy of Sciences108, 15727 (2011)
2011
-
[69]
Elgeti and G
J. Elgeti and G. Gompper, Emergence of metachronal waves in cilia arrays, Proceedings of the National Academy of Sciences110, 4470 (2013)
2013
-
[70]
J. R. Blake, A note on the image system for a stokeslet in a no-slip boundary, Mathematical Proceedings of the Cambridge Philosophical Society70, 303 (1971)
1971
-
[71]
J. W. Swan and J. F. Brady, Simulation of hydrodynamically interacting particles near a no-slip boundary, Physics of Fluids19(2007)
2007
-
[72]
Liron, Fluid transport by cilia between parallel plates, Journal of Fluid Mechanics86, 705 (1978)
N. Liron, Fluid transport by cilia between parallel plates, Journal of Fluid Mechanics86, 705 (1978)
1978
-
[73]
D. J. Smith, E. A. Gaffney, and J. R. Blake, Modelling mucociliary clearance, Respiratory physiology & neurobiology163, 178 (2008)
2008
-
[74]
Proximal Policy Optimization Algorithms
J. Schulman, F. Wolski, P. Dhariwal, A. Radford, and O. Klimov, Proximal policy optimization algorithms, arXiv preprint arXiv:1707.06347 (2017)
work page internal anchor Pith review Pith/arXiv arXiv 2017
-
[75]
R. S. Sutton, A. G. Barto,et al.,Reinforcement learning: An introduction, Vol. 1 (MIT press Cambridge, 1998)
1998
-
[76]
Eloy and E
C. Eloy and E. Lauga, Kinematics of the most efficient cilium, Physical Review Letters109, 038101 (2012)
2012
-
[77]
M. C. Lagomarsino, F. Capuani, and C. P. Lowe, A simulation study of the dynamics of a driven filament in an aristotelian fluid, Journal of theoretical biology224, 215 (2003)
2003
-
[78]
Khaderi, J
S. Khaderi, J. Den Toonder, and P. Onck, Fluid flow due to collective non-reciprocal motion of symmetrically-beating artificial cilia, Biomicrofluidics6(2012)
2012
-
[79]
T. Wang, T. ul Islam, E. Steur, T. Homan, I. Aggarwal, P. R. Onck, J. M. den Toonder, and Y. Wang, Programmable metachronal motion of closely packed magnetic artificial cilia, Lab on a Chip24, 1573 (2024)
2024
-
[80]
Uchida and R
N. Uchida and R. Golestanian, Hydrodynamic synchronization between objects with cyclic rigid trajectories, The European Physical Journal E35, 135 (2012)
2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.