DNA Replication under Thermal, Chemical, and Genotoxic Stress
Pith reviewed 2026-06-27 19:02 UTC · model grok-4.3
The pith
Replication fork speed heterogeneity produces Erlang-distributed S-phase durations and explains stress responses.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
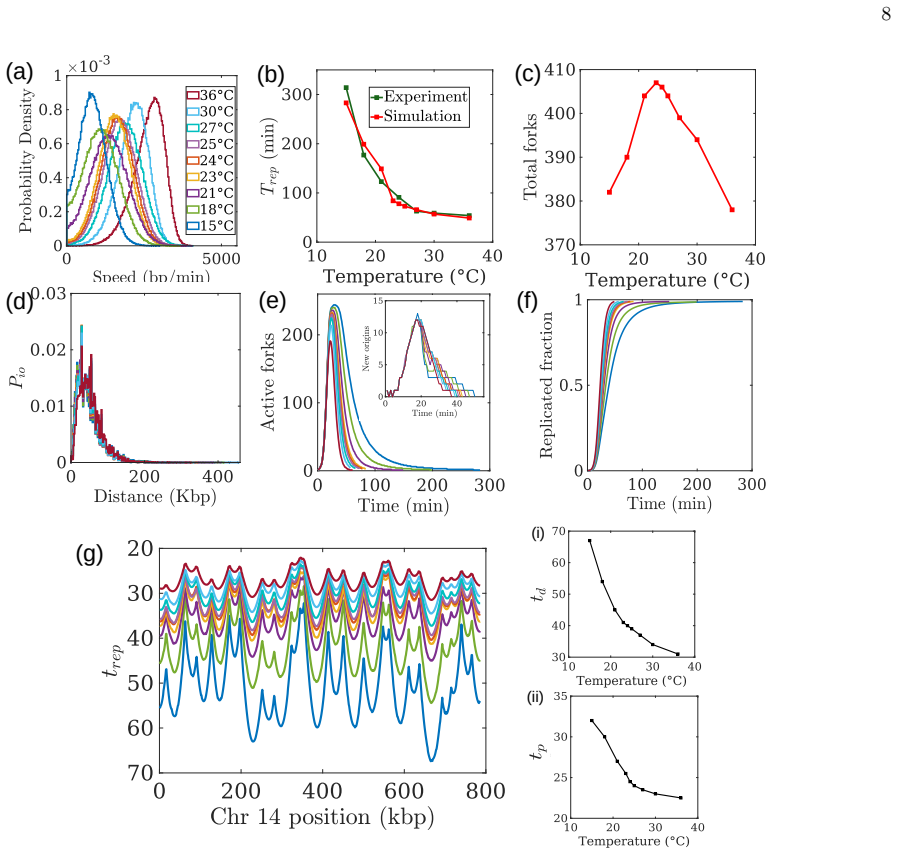
The central claim is that replication fork-speed heterogeneity is the underlying cause of Erlang-distributed S-phase durations and rare, anomalously prolonged replication events. The lattice-based stochastic Monte Carlo model, benchmarked against experimental replication profiles, reproduces these distributions and predicts non-monotonic thermal dependence, power-law scaling under hydroxyurea, and characteristic total replication times under diverse genotoxic conditions, all with only two effective parameters.
What carries the argument
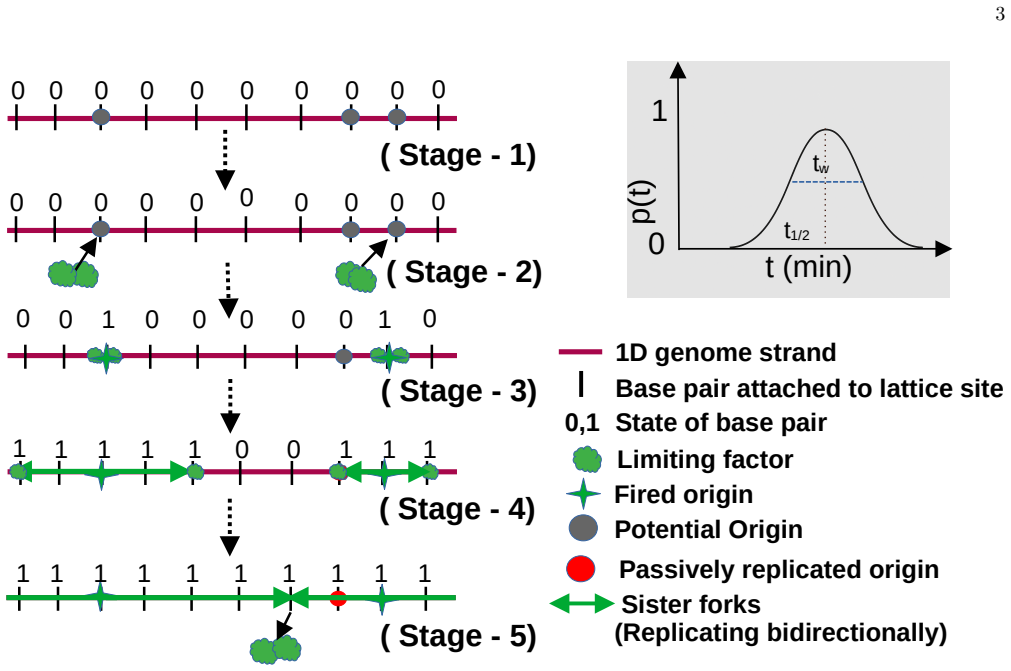
A lattice-based stochastic Monte Carlo framework at single base-pair resolution that incorporates probabilistic origin firing, replication fork-speed distributions, and a time-dependent limiting factor on cellular resources.
If this is right
- Fork-speed heterogeneity generates Erlang-distributed S-phase durations across organisms.
- Rare, anomalously long replication events appear in S. cerevisiae as they do in E. coli and human cells.
- Total replication time exhibits non-monotonic dependence on temperature.
- Replication dynamics follow power-law scaling under hydroxyurea stress.
- Distinct total replication-time patterns emerge under different genotoxic conditions.
Where Pith is reading between the lines
- The two-parameter structure implies that a common resource-limitation mechanism operates across unrelated stress types.
- Measuring fork-speed distributions directly in yeast under controlled thermal stress would test the non-monotonic prediction.
- The framework could be extended to mammalian genomes by changing only the origin-firing statistics while retaining the same fork heterogeneity.
- If fork-speed variation dominates timing, then mutations that narrow the speed distribution should reduce the frequency of long S-phase events.
Load-bearing premise
That a lattice-based Monte Carlo model using only two effective parameters can reproduce the full range of observed replication responses to thermal, chemical, and genotoxic stress when benchmarked against experimental profiles.
What would settle it
Experimental observation that S-phase durations in yeast do not follow an Erlang distribution or that replication times fail to show the predicted non-monotonic temperature dependence or power-law scaling under hydroxyurea.
Figures



read the original abstract
Eukaryotic DNA replication must remain robust under thermal, chemical, and genotoxic stress despite large fluctuations in replication dynamics. Here, we develop a lattice-based stochastic Monte Carlo framework for whole-genome replication in Saccharomyces cerevisiae at single base-pair resolution, incorporating probabilistic origin firing, replication fork-speed distributions, and a time-dependent limiting factor that governs the availability of cellular replication resources. The model is benchmarked quantitatively against experimental replication profiles before being applied to stress conditions, and reproduces diverse replication stress responses using only two effective parameters. Importantly, the analysis reveals that replication fork-speed heterogeneity underlies the emergence of Erlang-distributed S-phase durations and rare, anomalously prolonged replication events observed experimentally in Escherichia coli and human cell lines, while predicting similar behavior in S. cerevisiae. The framework further predicts non-monotonic thermal behavior, power-law scaling under hydroxyurea stress, and total replication-time dynamics under diverse genotoxic conditions.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper presents a lattice-based stochastic Monte Carlo model of whole-genome DNA replication in S. cerevisiae at single base-pair resolution. The framework incorporates probabilistic origin firing, replication fork-speed distributions, and a time-dependent limiting factor for cellular resources. It is benchmarked against experimental profiles and applied to thermal, chemical (hydroxyurea), and genotoxic stress, claiming to reproduce Erlang-distributed S-phase durations, rare prolonged events, non-monotonic thermal responses, power-law scaling under hydroxyurea, and total replication-time dynamics across conditions using only two effective parameters. The analysis attributes Erlang statistics and anomalous events to fork-speed heterogeneity and extends predictions to E. coli, human cells, and S. cerevisiae.
Significance. If the central claims hold with fixed parameters across regimes, the work would supply a mechanistic link between microscopic fork heterogeneity and observed S-phase statistics, offering a unified account of replication robustness under diverse stresses with falsifiable predictions for non-monotonic and power-law behaviors. The quantitative benchmarking and single-base-pair resolution are strengths, though the low parameter count requires verification that the reproduction is not achieved by per-condition adjustment.
major comments (2)
- [Abstract] Abstract and model description: the central claim that diverse stress responses are reproduced 'using only two effective parameters' is load-bearing; the manuscript must explicitly state and demonstrate whether these two parameters remain fixed across all thermal, chemical, and genotoxic regimes or are refit separately for each dataset. If refitting occurs, the reproduction reduces to a flexible phenomenological description rather than a unified mechanistic prediction.
- [Model] Model section (description of fork-speed input): because fork-speed distributions are supplied as direct input to the Monte Carlo lattice, the reported emergence of Erlang-distributed S-phase durations and rare prolonged events is at least partly by construction; the paper must quantify how much of the Erlang shape and tail statistics arise independently of the input distribution versus from the interaction with the limiting factor and origin firing.
minor comments (1)
- [Abstract] The abstract states quantitative benchmarking against experimental profiles but provides no details on error metrics, cross-validation, or independent test datasets; these should be added for clarity.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive report. The comments identify important points for clarification regarding parameter usage and the origins of the reported statistical behaviors. We address each major comment below and have revised the manuscript to incorporate the requested explicit statements and additional analyses.
read point-by-point responses
-
Referee: [Abstract] Abstract and model description: the central claim that diverse stress responses are reproduced 'using only two effective parameters' is load-bearing; the manuscript must explicitly state and demonstrate whether these two parameters remain fixed across all thermal, chemical, and genotoxic regimes or are refit separately for each dataset. If refitting occurs, the reproduction reduces to a flexible phenomenological description rather than a unified mechanistic prediction.
Authors: We agree that explicit demonstration is required. The two effective parameters (the resource-limiting factor scaling coefficient and the global origin-firing rate prefactor) are held fixed at the values determined from the unstressed benchmark across all thermal, hydroxyurea, and genotoxic conditions. Stress enters only through the measured or modeled input distributions for fork speeds and, where relevant, origin efficiencies. In the revised manuscript we have added a dedicated paragraph in the Model section, an updated abstract sentence, and a new supplementary table that lists the identical numerical values of these two parameters for every regime together with a brief sensitivity test confirming that independent refitting is not performed. revision: yes
-
Referee: [Model] Model section (description of fork-speed input): because fork-speed distributions are supplied as direct input to the Monte Carlo lattice, the reported emergence of Erlang-distributed S-phase durations and rare prolonged events is at least partly by construction; the paper must quantify how much of the Erlang shape and tail statistics arise independently of the input distribution versus from the interaction with the limiting factor and origin firing.
Authors: We acknowledge that the input distribution contributes to the output. Nevertheless, the characteristic Erlang shape and the heavy tail of prolonged events are generated by the stochastic superposition of many independent forks whose instantaneous speeds are drawn from the distribution, combined with the time-dependent limiting factor and probabilistic origin firing. To quantify the separate contributions we have performed additional control simulations (now reported in the revised Supplementary Information) in which the heterogeneous fork-speed distribution is replaced by a constant speed equal to its mean while every other model component remains unchanged. These controls show that both the Erlang shape parameter and the probability of extreme tails are substantially degraded, thereby demonstrating that the interaction with the limiting factor and origin dynamics is essential. The main text has been updated with a concise statement of this result and the associated figure reference. revision: yes
Axiom & Free-Parameter Ledger
free parameters (1)
- two effective parameters
axioms (1)
- domain assumption Lattice-based stochastic Monte Carlo simulation is a valid representation of eukaryotic DNA replication dynamics
Reference graph
Works this paper leans on
-
[1]
The fork speed for this mutant shows a much narrower distribution with similar mean fork speed as compared to M0 (Fig. 6a). This might be indicative of the possibility that forks that were earlier progressing slowly start replicating much faster owing to the higher availability of dNTPs. This also allows slightly early completion of DNA replication, as sh...
2022
-
[2]
P. Zhou, X. Gao, X. Li, L. Li, C. Niu, Q. Ouyang, H. Lou, T. Li, and F. Li, Stochasticity triggers activation of the s-phase checkpoint pathway in budding yeast, Physical Review X11, 011004 (2021)
2021
-
[3]
Willis and N
N. Willis and N. Rhind, Regulation of dna replication by the s-phase dna damage checkpoint, Cell division4, 13 (2009)
2009
-
[4]
Gaillard, T
H. Gaillard, T. Garc´ ıa-Muse, and A. Aguilera, Replication stress and cancer, Nature Reviews Cancer15, 276 (2015)
2015
-
[5]
M. D. Sekedat, D. Feny¨ o, R. S. Rogers, A. J. Tackett, J. D. Aitchison, and B. T. Chait, Gins motion reveals replication fork progression is remarkably uniform throughout the yeast genome, Molecular systems biology6, 353 (2010)
2010
-
[6]
Berners-Lee, E
R. Berners-Lee, E. Gilmore, F. Berkemeier, and M. A. Boemo, Regulation of replication timing in saccharomyces cerevisiae, PLOS Computational Biology21, e1013066 (2025)
2025
-
[7]
Goffeau, B
A. Goffeau, B. G. Barrell, H. Bussey, R. W. Davis, B. Dujon, H. Feldmann, F. Galibert, J. D. Hoheisel, C. Jacq, M. John- ston,et al., Life with 6000 genes, Science274, 546 (1996). 12
1996
-
[8]
A. M. Breier, S. Chatterji, and N. R. Cozzarelli, Prediction of saccharomyces cerevisiae replication origins, Genome biology 5, R22 (2004)
2004
-
[9]
Chang, C
F. Chang, C. D. May, T. Hoggard, J. Miller, C. A. Fox, and M. Weinreich, High-resolution analysis of four efficient yeast replication origins reveals new insights into the orc and putative mcm binding elements, Nucleic acids research39, 6523 (2011)
2011
-
[10]
Fosset al., Identification of 1600 replication origins in s
E. Fosset al., Identification of 1600 replication origins in s. cerevisiae. elife 12, RP880871025, 1026 (2024)
2024
-
[11]
Dukaj and N
L. Dukaj and N. Rhind, The capacity of origins to load mcm establishes replication timing patterns, PLoS Genetics17, e1009467 (2021)
2021
-
[12]
Labib, How do cdc7 and cyclin-dependent kinases trigger the initiation of chromosome replication in eukaryotic cells?, Genes & development24, 1208 (2010)
K. Labib, How do cdc7 and cyclin-dependent kinases trigger the initiation of chromosome replication in eukaryotic cells?, Genes & development24, 1208 (2010)
2010
-
[13]
De Jes´ us-Kim, L
L. De Jes´ us-Kim, L. J. Friedman, M. L˜ ooke, C. K. Ramsoomair, J. Gelles, and S. P. Bell, Ddk regulates replication initiation by controlling the multiplicity of cdc45-gins binding to mcm2-7, Elife10, e65471 (2021)
2021
-
[14]
K¨ ohler, D
C. K¨ ohler, D. Koalick, A. Fabricius, A. C. Parplys, K. Borgmann, H. Pospiech, and F. Grosse, Cdc45 is limiting for replication initiation in humans, Cell Cycle15, 974 (2016)
2016
-
[15]
Z. Yuan, R. Georgescu, L. Bai, D. Zhang, H. Li, and M. E. O’Donnell, Dna unwinding mechanism of a eukaryotic replicative cmg helicase, Nature communications11, 688 (2020)
2020
-
[16]
Rhind, Dna replication timing: Biochemical mechanisms and biological significance, Bioessays44, 2200097 (2022)
N. Rhind, Dna replication timing: Biochemical mechanisms and biological significance, Bioessays44, 2200097 (2022)
2022
-
[17]
P. K. Patel, N. Kommajosyula, A. Rosebrock, A. Bensimon, J. Leatherwood, J. Bechhoefer, and N. Rhind, The hsk1 (cdc7) replication kinase regulates origin efficiency, Molecular biology of the cell19, 5550 (2008)
2008
-
[18]
Mantiero, A
D. Mantiero, A. Mackenzie, A. Donaldson, and P. Zegerman, Limiting replication initiation factors execute the temporal programme of origin firing in budding yeast, The EMBO journal30, 4805 (2011)
2011
-
[19]
Y. Xia, The fate of two unstoppable trains after arriving destination: replisome disassembly during dna replication termi- nation, Frontiers in cell and developmental biology9, 658003 (2021)
2021
-
[20]
L¨ ob, N
D. L¨ ob, N. Lengert, V. Chagin, M. Reinhart, C. Casas-Delucchi, M. Cardoso, and B. Drossel, 3d replicon distributions arise from stochastic initiation and domino-like dna replication progression, Nature communications7, 11207 (2016)
2016
-
[21]
Theulot, L
B. Theulot, L. Lacroix, J.-M. Arbona, G. A. Millot, E. Jean, C. Cruaud, J. Pellet, F. Proux, M. Hennion, S. Engelen, et al., Genome-wide mapping of individual replication fork velocities using nanopore sequencing, Nature communications 13, 3295 (2022)
2022
-
[22]
Azvolinsky, P
A. Azvolinsky, P. G. Giresi, J. D. Lieb, and V. A. Zakian, Highly transcribed rna polymerase ii genes are impediments to replication fork progression in saccharomyces cerevisiae, Molecular cell34, 722 (2009)
2009
-
[23]
Pellet, L
J. Pellet, L. Lacroix, B. Theulot, E. Simonin Chavignier, A. Tourancheau, F. Proux, S. Hermann-Le Denmat, G. A. Millot, B. Le Tallec, and O. Hyrien, Replication program of a single-chromosome budding yeast strain, Nucleic Acids Research 53, gkaf754 (2025)
2025
-
[24]
Labit, I
H. Labit, I. Perewoska, T. Germe, O. Hyrien, and K. Marheineke, Dna replication timing is deterministic at the level of chromosomal domains but stochastic at the level of replicons in xenopus egg extracts, Nucleic acids research36, 5623 (2008)
2008
-
[25]
Hawkins, R
M. Hawkins, R. Retkute, C. A. M¨ uller, N. Saner, T. U. Tanaka, A. P. de Moura, and C. A. Nieduszynski, High-resolution replication profiles define the stochastic nature of genome replication initiation and termination, Cell reports5, 1132 (2013)
2013
-
[26]
D. M. Gilbert, Evaluating genome-scale approaches to eukaryotic dna replication, Nature Reviews Genetics11, 673 (2010)
2010
-
[27]
Barberis, T
M. Barberis, T. W. Spiesser, and E. Klipp, Replication origins and timing of temporal replication in budding yeast: how to solve the conundrum?, Changing Views on Living Organisms , 15 (2013)
2013
-
[28]
K. L. Friedman, B. J. Brewer, and W. L. Fangman, Replication profile of saccharomyces cerevisiae chromosome vi, Genes to Cells2, 667 (1997)
1997
-
[29]
Tuduri, H
S. Tuduri, H. Tourri` ere, and P. Pasero, Defining replication origin efficiency using dna fiber assays, Chromosome Research 18, 91 (2010)
2010
-
[30]
Kelly and A
T. Kelly and A. J. Callegari, Dynamics of dna replication in a eukaryotic cell, Proceedings of the National Academy of Sciences116, 4973 (2019)
2019
-
[31]
P. K. Patel, B. Arcangioli, S. P. Baker, A. Bensimon, and N. Rhind, Dna replication origins fire stochastically in fission yeast, Molecular biology of the cell17, 308 (2006)
2006
-
[32]
Vanoni, M
M. Vanoni, M. Vai, and G. Frascotti, Effects of temperature on the yeast cell cycle analyzed by flow cytometry, Cytometry: The Journal of the International Society for Analytical Cytology5, 530 (1984)
1984
-
[33]
J. Poli, O. Tsaponina, L. Crabb´ e, A. Keszthelyi, V. Pantesco, A. Chabes, A. Lengronne, and P. Pasero, dntp pools determine fork progression and origin usage under replication stress, The EMBO journal31, 883 (2012)
2012
-
[34]
Rhind, Dna replication timing: random thoughts about origin firing, Nature cell biology8, 1313 (2006)
N. Rhind, Dna replication timing: random thoughts about origin firing, Nature cell biology8, 1313 (2006)
2006
-
[35]
Berkemeier, P
F. Berkemeier, P. R. Cook, and M. A. Boemo, Dna replication timing reveals genome-wide features of transcription and fragility, Nature Communications16, 4658 (2025)
2025
-
[36]
E. Ma, O. Hyrien, and A. Goldar, Do replication forks control late origin firing in saccharomyces cerevisiae?, Nucleic acids research40, 2010 (2012)
2010
-
[37]
R. Pugatch, Greedy scheduling of cellular self-replication leads to optimal doubling times with a log-frechet distribution, Proceedings of the National Academy of Sciences112, 2611 (2015)
2015
-
[38]
H. X. Chao, R. I. Fakhreddin, H. K. Shimerov, K. M. Kedziora, R. J. Kumar, J. Perez, J. C. Limas, G. D. Grant, J. G. Cook, G. P. Gupta,et al., Evidence that the human cell cycle is a series of uncoupled, memoryless phases, Molecular systems biology15, MSB188604 (2019). 13
2019
-
[39]
A. E. Shaw, M. N. Mihelich, J. E. Whitted, H. J. Reitman, A. J. Timmerman, M. Tehseen, S. M. Hamdan, and G. D. Schauer, Revised mechanism of hydroxyurea-induced cell cycle arrest and an improved alternative, Proceedings of the National Academy of Sciences121, e2404470121 (2024)
2024
-
[40]
Kumar, J
D. Kumar, J. Viberg, A. K. Nilsson, and A. Chabes, Highly mutagenic and severely imbalanced dntp pools can escape detection by the s-phase checkpoint, Nucleic acids research38, 3975 (2010)
2010
-
[41]
G. M. Alvino, D. Collingwood, J. M. Murphy, J. Delrow, B. J. Brewer, and M. Raghuraman, Replication in hydroxyurea: it’s a matter of time, Molecular and cellular biology27, 6396 (2007)
2007
-
[42]
Frenkel, F
N. Frenkel, F. Jonas, M. Carmi, G. Yaakov, and N. Barkai, Rtt109 slows replication speed by histone n-terminal acetylation, Genome research31, 426 (2021)
2021
-
[43]
Kaplan, C
T. Kaplan, C. L. Liu, J. A. Erkmann, J. Holik, M. Grunstein, P. D. Kaufman, N. Friedman, and O. J. Rando, Cell cycle–and chaperone-mediated regulation of h3k56ac incorporation in yeast, PLoS genetics4, e1000270 (2008)
2008
-
[44]
S. Ide, K. Saka, and T. Kobayashi, Rtt109 prevents hyper-amplification of ribosomal rna genes through histone modification in budding yeast, PLoS genetics9, e1003410 (2013)
2013
-
[45]
J. Han, H. Zhou, Z. Li, R.-M. Xu, and Z. Zhang, Acetylation of lysine 56 of histone h3 catalyzed by rtt109 and regulated by asf1 is required for replisome integrity, Journal of Biological Chemistry282, 28587 (2007)
2007
-
[46]
X. Zhao, A. Chabes, V. Domkin, L. Thelander, and R. Rothstein, The ribonucleotide reductase inhibitor sml1 is a new target of the mec1/rad53 kinase cascade during growth and in response to dna damage, The EMBO journal (2001)
2001
-
[47]
J. T. Yeeles, A. Janska, A. Early, and J. F. Diffley, How the eukaryotic replisome achieves rapid and efficient dna replication, Molecular cell65, 105 (2017)
2017
-
[48]
Tourri` ere, G
H. Tourri` ere, G. Versini, V. Cord´ on-Preciado, C. Alabert, and P. Pasero, Mrc1 and tof1 promote replication fork progression and recovery independently of rad53, Molecular cell19, 699 (2005). S1 SUPPLEMENT AR Y INFORMA TION (d) (a) (b) (c) FIG. S1. (a) Time evolution of fired-origin intensity. (b) Activation functionA f, defined as the ratio of the num...
2005
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.