Spontaneous polarization for protrusion-driven cell crawling
Pith reviewed 2026-06-27 10:48 UTC · model grok-4.3
The pith
Feedback between cell motion and polarized actin nucleation regulator drives spontaneous crawling above a critical activity threshold.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
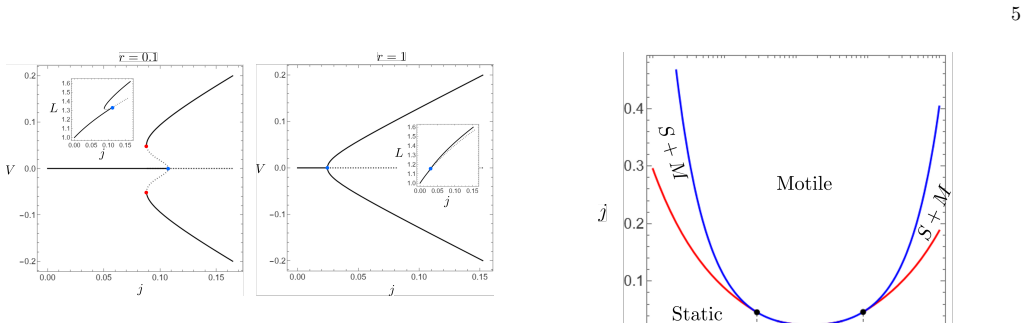
In the minimal one-dimensional model the static symmetric cell state loses stability above a critical protrusive activity through a bifurcation induced by the feedback between boundary motion and polarized chemical regulation of actin nucleation, resulting in spontaneous transition to a motile state with unequal polymerization rates at the two edges.
What carries the argument
The feedback loop in which cell motion polarizes an external chemical regulator of actin nucleation, thereby imposing different polymerization densities at the two moving edges.
Load-bearing premise
The external chemical regulator of actin nucleation becomes polarized around the moving cell boundaries and thereby imposes different nucleation densities at the two edges.
What would settle it
Direct measurement showing whether the external regulator concentration differs measurably between the leading and trailing edges in a spontaneously moving cell, or experimental tuning of protrusive activity across the model's predicted critical value to test for the onset of motility.
Figures

read the original abstract
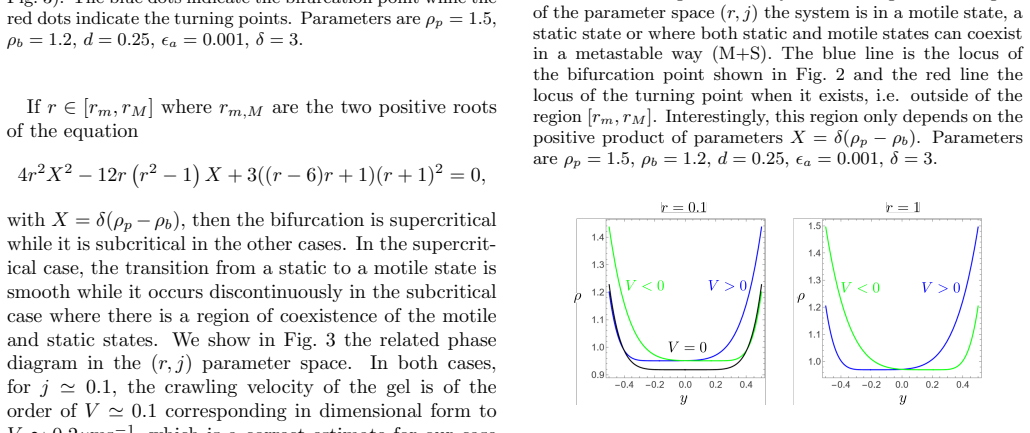
We propose a minimal one-dimensional continuum model for the spontaneous initiation of protrusion-driven cell crawling on a rigid substrate. The cell cytoskeleton is represented as a viscous actin meshwork that turns over in the bulk and polymerizes at two moving cell edges. Symmetry breaking arises from the feedback between cell motion, an external chemical regulator of actin nucleation, and actin polymerization at the cell fronts. When the cell moves, the regulator becomes polarized around the moving boundaries, thereby imposing different actin nucleation densities at the two edges. This generates unequal protrusive rates, which in turn reinforce motion and sustain the chemical polarization. Above a critical protrusive activity, the static symmetric state loses stability and the system undergoes a bifurcation toward a motile polarized state. Depending on how the external cue controls actin nucleation, the transition can be either supercritical or subcritical, leading in the latter case to coexistence between static and motile states. Using parameter values appropriate for keratocyte cells, the model predicts realistic crawling speeds and actin-density profiles, including asymmetric edge-localized density peaks. These results identify a generic mechanism by which external biochemical regulation of actin nucleation can trigger spontaneous motility along a one-dimensional track without requiring molecular motors, specific adhesion dynamics, deformable substrates, or pre-existing polarity.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes a minimal one-dimensional continuum model for protrusion-driven cell crawling on a rigid substrate. The cytoskeleton is modeled as a viscous actin meshwork with bulk turnover and edge polymerization; symmetry breaking arises from positive feedback between cell velocity, polarization of an external chemical regulator of actin nucleation, and differential protrusive activity at the two edges. Above a critical protrusive activity the symmetric static state loses stability via a bifurcation to a motile polarized state; the bifurcation is supercritical or subcritical depending on the functional form of nucleation regulation. With keratocyte-appropriate parameters the model yields realistic crawling speeds and asymmetric actin-density profiles.
Significance. If the derivation and stability analysis hold, the work identifies a generic, motor-independent mechanism for spontaneous motility initiation driven by external biochemical regulation. The demonstration that the same feedback loop can produce either continuous or hysteretic transitions is biologically relevant for understanding bistability in cell polarity. The 1D continuum formulation is analytically tractable and reproduces keratocyte-scale outputs, providing a useful minimal model for further exploration.
major comments (2)
- [Model description / abstract paragraph on feedback loop] The abstract states that motion polarizes the external regulator around the moving boundaries, but neither the abstract nor the model description supplies the explicit transport or boundary condition that implements this polarization (e.g., an advection term proportional to velocity or a flux condition at the edges). Without this equation the feedback loop cannot be verified as emergent rather than imposed, which is load-bearing for the claimed spontaneous symmetry breaking.
- [Results paragraph on keratocyte parameters] Parameter values are chosen to match keratocyte data, yet no sensitivity analysis or cross-validation is reported; it is therefore unclear whether the predicted speeds and density profiles are independent outputs or post-fit results. This directly affects the strength of the claim that the model reproduces realistic crawling without additional mechanisms.
minor comments (2)
- The abstract mentions 'model equations' but does not display them; the full manuscript should include the complete set of PDEs, boundary conditions, and the explicit functional form of the nucleation regulation that controls the super- versus subcritical character.
- Stability analysis details (linearization around the symmetric state, eigenvalue calculation, and bifurcation normal-form coefficients) are referenced only by outcome; these steps should be shown explicitly, preferably with the critical threshold expressed in terms of the model parameters.
Simulated Author's Rebuttal
We thank the referee for their constructive comments, which help clarify the presentation of the feedback mechanism and strengthen the robustness of our results. We address each point below and will revise the manuscript to improve explicitness and add supporting analysis.
read point-by-point responses
-
Referee: [Model description / abstract paragraph on feedback loop] The abstract states that motion polarizes the external regulator around the moving boundaries, but neither the abstract nor the model description supplies the explicit transport or boundary condition that implements this polarization (e.g., an advection term proportional to velocity or a flux condition at the edges). Without this equation the feedback loop cannot be verified as emergent rather than imposed, which is load-bearing for the claimed spontaneous symmetry breaking.
Authors: We agree that the explicit transport equation was not stated with sufficient prominence. The model (Section 2) implements polarization via the advection term in the regulator equation, δ_t c + v δ_x c = D δ_xx c - γ c, together with no-flux conditions at the moving edges x = ± L(t)/2. This advection arises directly from cell velocity v and produces the polarization without being imposed by hand. To make the loop fully verifiable, we will revise the abstract and expand the model-description paragraph to quote the transport equation and boundary conditions explicitly. revision: yes
-
Referee: [Results paragraph on keratocyte parameters] Parameter values are chosen to match keratocyte data, yet no sensitivity analysis or cross-validation is reported; it is therefore unclear whether the predicted speeds and density profiles are independent outputs or post-fit results. This directly affects the strength of the claim that the model reproduces realistic crawling without additional mechanisms.
Authors: We accept that the absence of sensitivity analysis weakens the claim of independent reproduction. We will add a new subsection (or appendix) reporting a systematic sensitivity sweep over the biologically relevant ranges of protrusive activity, regulator diffusivity, and turnover rates. The analysis will show that realistic speeds (0.1–0.5 μm s^{-1}) and edge-density peaks persist across wide intervals, confirming the outputs are robust rather than narrowly tuned. revision: yes
Circularity Check
No significant circularity; derivation is self-contained bifurcation analysis
full rationale
The central result is a standard linear stability analysis of the symmetric equilibrium in a minimal 1D continuum model, with the bifurcation type (super- or subcritical) controlled by the functional form of the nucleation regulation term. No quoted step reduces a claimed prediction to a fitted parameter by construction, nor does any load-bearing premise rest on a self-citation chain. Parameter selection for keratocyte scales is used only to generate illustrative outputs after the bifurcation threshold is derived; the symmetry-breaking mechanism itself follows directly from the stated feedback equations without circular reduction. The paper is therefore scored at the default non-circular level.
Axiom & Free-Parameter Ledger
free parameters (2)
- critical protrusive activity threshold
- keratocyte-specific nucleation and turnover rates
axioms (2)
- domain assumption Actin cytoskeleton behaves as a one-dimensional viscous meshwork with bulk turnover and edge polymerization only.
- domain assumption External chemical regulator polarization is strictly determined by cell motion.
invented entities (1)
-
external chemical regulator of actin nucleation
no independent evidence
Reference graph
Works this paper leans on
-
[1]
activated
0.25 Morphogen turnoverϵ a =η/(kτ a) 10 −3 Actin turnoverϵ=η/(kτ) 2.5 TABLE I. Estimates of material coefficients (case of kerato- cyte cells) and nondimensional parameter definitions. nondimensional model reads: ∂s(ρL)−∂ y(ρ[L(L−1)y+ ˙C+y ˙L]) =ϵL(¯ρ−ρ) ∂s(aL)−∂ y(a( ˙C+y ˙L) + d L ∂ya) =ϵ aL(1−a) ∓jexp(δ(ρ| ∓1/2 −1)) = ˙l∓ ∓L(L−1)/2 d L ∂ya|∓1/2 +a| ∓1/...
-
[2]
BothL(t) andV(t) are scalar parameters that need to be solved for along with the fieldsρ(y, t) (fory∈[−1/2,1/2]) anda(y, t) (fory∈]− ∞,−1/2[∪]1/2,∞[)
represents the diffusion of the morphogena,r=a 0/a∗ is the ratio between the equi- librium concentration of the morphogen outside the cell and the threshold concentration upon which it switches the nucleation density of actin,ϵ=η/(kτ) is the bulk turnover rate of actin andϵ a =η/(kτ a) is the typical degradation rate of the morphogen. BothL(t) andV(t) are...
-
[3]
However, it is unclear in our case why Arpin would be advected with the actin meshwork in normal conditions
whereais associated with Arpin is to consider thata is advected with the actin meshwork and follows ∂ta+∂ x(av)−D∂ xxa= 0.(16) If we make this change in our model, we recover an insta- bility of the same type as the one described here because a drift remains in (16) even in the co-moving frame since vandVare different. However, it is unclear in our case w...
-
[4]
de Beco, K
S. de Beco, K. Vaidˇ ziulyt˙ e, J. Manzi, F. Dalier, F. Di Fed- erico, G. Cornilleau, M. Dahan, and M. Coppey, Nature communications9, 4816 (2018)
2018
-
[5]
S. K. Raja, M. S. Garcia, R. R. Isseroff,et al., Front Biosci12, 2849 (2007)
2007
-
[6]
R. E. Roberts and M. B. Hallett, International journal of molecular sciences20, 1383 (2019)
2019
-
[7]
A. C. Callan-Jones and R. Voituriez, Current opinion in cell biology38, 12 (2016)
2016
-
[8]
Barnhart, K.-C
E. Barnhart, K.-C. Lee, G. M. Allen, J. A. Theriot, and A. Mogilner, Proceedings of the National Academy of Sciences112, 5045 (2015)
2015
-
[9]
Sens, Proceedings of the National Academy of Sciences 117, 24670 (2020)
P. Sens, Proceedings of the National Academy of Sciences 117, 24670 (2020)
2020
-
[10]
J. E. Ron, P. Monzo, N. C. Gauthier, R. Voituriez, and N. S. Gov, Physical review research2, 033237 (2020)
2020
-
[11]
Recho, T
P. Recho, T. Putelat, and L. Truskinovsky, Physical re- view letters111, 108102 (2013)
2013
-
[12]
Callan-Jones and R
A. Callan-Jones and R. Voituriez, New Journal of Physics 15, 025022 (2013)
2013
-
[13]
I. Lavi, N. Meunier, R. Voituriez, and J. Casademunt, Physical Review E101, 022404 (2020)
2020
-
[14]
N. O. Winkler, O. M. Drozdowski, F. Ziebert, and U. S. Schwarz, Physical Review E112, 024413 (2025)
2025
-
[15]
Ziebert, S
F. Ziebert, S. Swaminathan, and I. S. Aranson, Journal of The Royal Society Interface9, 1084 (2012)
2012
-
[16]
B. A. Camley, Y. Zhao, B. Li, H. Levine, and W.-J. Rap- pel, Physical review letters111, 158102 (2013)
2013
-
[17]
Blanch-Mercader and J
C. Blanch-Mercader and J. Casademunt, Physical review letters110, 078102 (2013)
2013
-
[18]
Schmidt, W
W. Schmidt, W. Zimmermann, C. Misbah, and A. Farutin, Physical Review Letters134, 158301 (2025)
2025
-
[19]
Tjhung, D
E. Tjhung, D. Marenduzzo, and M. E. Cates, Proceedings of the National Academy of Sciences109, 12381 (2012)
2012
-
[20]
Giomi and A
L. Giomi and A. DeSimone, Physical review letters112, 147802 (2014)
2014
-
[21]
Y. Mori, A. Jilkine, and L. Edelstein-Keshet, Biophysical journal94, 3684 (2008)
2008
-
[22]
Vanderlei, J
B. Vanderlei, J. J. Feng, and L. Edelstein-Keshet, Mul- tiscale Modeling & Simulation9, 1420 (2011)
2011
-
[23]
Copos and A
C. Copos and A. Mogilner, Molecular biology of the cell 31, 1637 (2020)
2020
-
[24]
T. D. Pollard, W. C. Earnshaw, J. Lippincott-Schwartz, and G. T. Johnson,Cell Biology, 3rd ed. (Elsevier, Philadelphia, 2017)
2017
-
[25]
Mogilner and G
A. Mogilner and G. Oster, Biophysical journal71, 3030 (1996)
1996
-
[26]
C. S. Peskin, G. M. Odell, and G. F. Oster, Biophysical journal65, 316 (1993)
1993
-
[27]
Larripa and A
K. Larripa and A. Mogilner, Physica A: Statistical Me- chanics and its Applications372, 113 (2006)
2006
-
[28]
Julicher, K
F. Julicher, K. Kruse, J. Prost, and J.-F. Joanny, Physics reports449, 3 (2007)
2007
-
[29]
Recho and L
P. Recho and L. Truskinovsky, Mathematics and Mechan- ics of Solids21, 263 (2016)
2016
-
[30]
Abeyaratne and P
R. Abeyaratne and P. K. Purohit, Journal of the Royal Society Interface21(2024)
2024
-
[31]
Ambrosi and A
D. Ambrosi and A. Zanzottera, Physica D: Nonlinear Phenomena330, 58 (2016)
2016
-
[32]
Giverso and L
C. Giverso and L. Preziosi, Physical Review E98, 062402 (2018)
2018
-
[33]
Prost, J.-F
J. Prost, J.-F. Joanny, P. Lenz, and C. Sykes, inCell Motility(Springer, 2008) pp. 1–30
2008
-
[34]
van der Gucht, E
J. van der Gucht, E. Paluch, J. Plastino, and C. Sykes, Proceedings of the National Academy of Sciences102, 7847 (2005)
2005
-
[35]
K. John, P. Peyla, K. Kassner, J. Prost, and C. Misbah, 9 Physical review letters100, 068101 (2008)
2008
-
[36]
´Etienne and P
J. ´Etienne and P. Recho, Journal of the Mechanics and Physics of Solids183, 105526 (2024)
2024
-
[37]
Michelin, E
S. Michelin, E. Lauga, and D. Bartolo, Physics of Fluids 25(2013)
2013
-
[38]
J. L. Moran and J. D. Posner, Annual Review of Fluid Mechanics49, 511 (2017)
2017
-
[39]
Kruse, J.-F
K. Kruse, J.-F. Joanny, F. J¨ ulicher, J. Prost, and K. Seki- moto, The European Physical Journal E16, 5 (2005)
2005
-
[40]
Recho and L
P. Recho and L. Truskinovsky, Physical Review E—Statistical, Nonlinear, and Soft Matter Physics87, 022720 (2013)
2013
-
[41]
Putelat, P
T. Putelat, P. Recho, and L. Truskinovsky, Physical Re- view E97, 012410 (2018)
2018
-
[42]
Tawada and K
K. Tawada and K. Sekimoto, Journal of theoretical biol- ogy150, 193 (1991)
1991
-
[43]
Chelly and P
H. Chelly and P. Recho, Physical Review E105, 064401 (2022)
2022
-
[44]
Vallotton, G
P. Vallotton, G. Danuser, S. Bohnet, J.-J. Meister, and A. B. Verkhovsky, Molecular biology of the cell16, 1223 (2005)
2005
-
[45]
Chelly, A
H. Chelly, A. Jahangiri, M. Mireux, J. ´Etienne, D. K. Dysthe, C. Verdier, and P. Recho, International Journal of Non-Linear Mechanics139, 103897 (2022)
2022
-
[46]
Mogilner and L
A. Mogilner and L. Edelstein-Keshet, Biophysical journal 83, 1237 (2002)
2002
-
[47]
Godlewski and P.-A
E. Godlewski and P.-A. Raviart,Hyperbolic Systems of Conservation Laws, Math´ ematiques & Applications, Vol. 3–4 (Ellipses, Paris, 1991) p. 252
1991
-
[48]
T. M. Svitkina and G. G. Borisy, The Journal of cell biology145, 1009 (1999)
1999
-
[49]
Plastino, I
J. Plastino, I. Lelidis, J. Prost, and C. Sykes, European Biophysics Journal33, 310 (2004)
2004
-
[50]
Colin, T
A. Colin, T. Kotila, C. Gu´ erin, M. Orhant-Prioux, B. Vianay, A. Mogilner, P. Lappalainen, M. Th´ ery, and L. Blanchoin, The EMBO journal42, EMBJ2022112717 (2023)
2023
-
[51]
D. R.-B. Aroush, N. Ofer, E. Abu-Shah, J. Allard, O. Krichevsky, A. Mogilner, and K. Keren, Current Bi- ology27, 2963 (2017)
2017
-
[52]
Rubinstein, M
B. Rubinstein, M. F. Fournier, K. Jacobson, A. B. Verkhovsky, and A. Mogilner, Biophysical journal97, 1853 (2009)
2009
-
[53]
Recho, A
P. Recho, A. Hallou, and E. Hannezo, Proceedings of the National Academy of Sciences116, 5344 (2019)
2019
-
[54]
B. J. Rollins, T. M. O’Connell, G. Bennett, L. E. Burton, C. D. Stiles, and J. G. Rheinwald, Journal of cellular physiology139, 455 (1989)
1989
-
[55]
Supplementary material, mathematica file to be ap- pended
-
[56]
Maiuri, J.-F
P. Maiuri, J.-F. Rupprecht, S. Wieser, V. Ruprecht, O. B´ enichou, N. Carpi, M. Coppey, S. De Beco, N. Gov, C.-P. Heisenberg,et al., Cell161, 374 (2015)
2015
-
[57]
Bohnet, R
S. Bohnet, R. Ananthakrishnan, A. Mogilner, J.-J. Meis- ter, and A. B. Verkhovsky, Biophysical journal90, 1810 (2006)
2006
-
[58]
Recho and L
P. Recho and L. Truskinovsky, Physical Review E110, 054413 (2024)
2024
-
[59]
S. R. De Groot and P. Mazur,Non-equilibrium thermo- dynamics(Courier Corporation, 2013)
2013
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.