Aggregation as a Double-Edged Sword: Fear, Allee Effects, and Finite-Time Collapse
Pith reviewed 2026-06-26 22:10 UTC · model grok-4.3
The pith
Prey aggregation accelerates finite-time extinction in a disease-structured model with fear and Allee effects.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
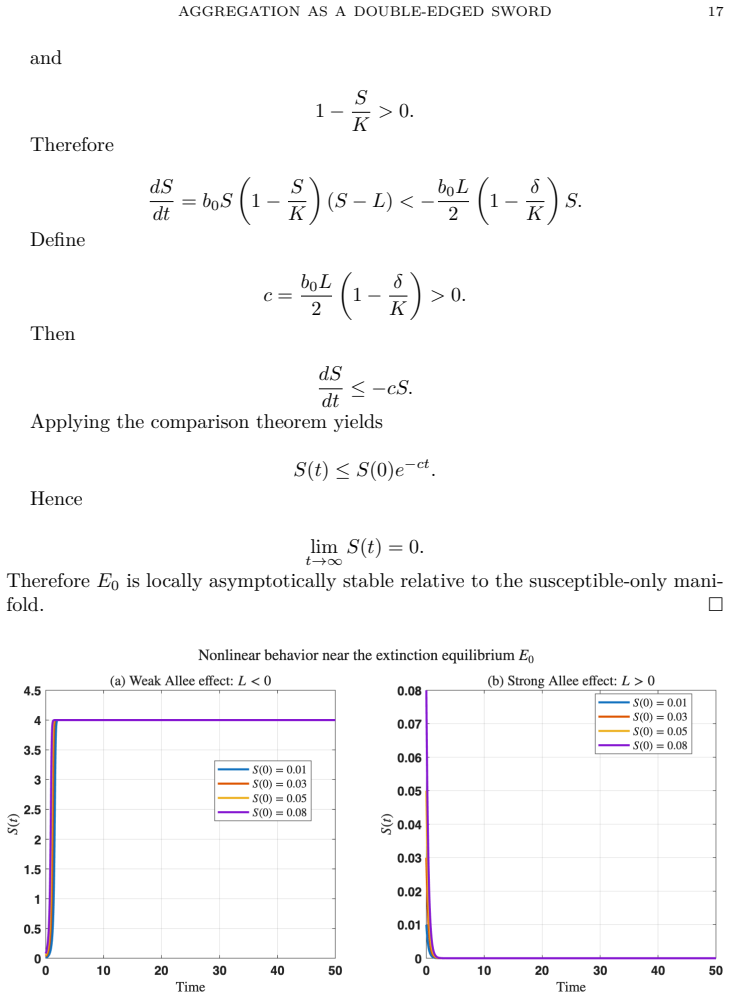
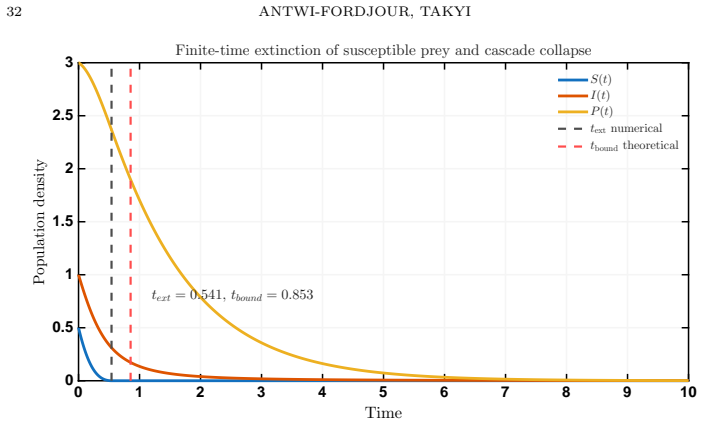
In the model, stronger aggregation monotonically enlarges the finite-time extinction region while providing a quantitative upper bound on extinction time that decreases with aggregation strength and predator pressure; this finite-time extinction of susceptible prey then triggers cascade collapse of the infected prey and predator populations.
What carries the argument
The explicit upper bound on extinction time, derived from the model equations, that quantifies how behavioral aggregation and demographic Allee thresholds jointly set the speed of ecological collapse.
If this is right
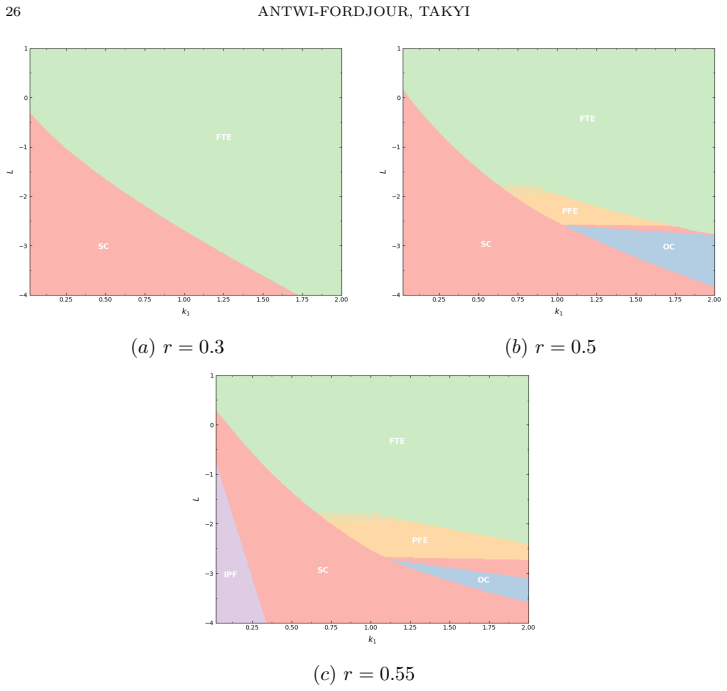
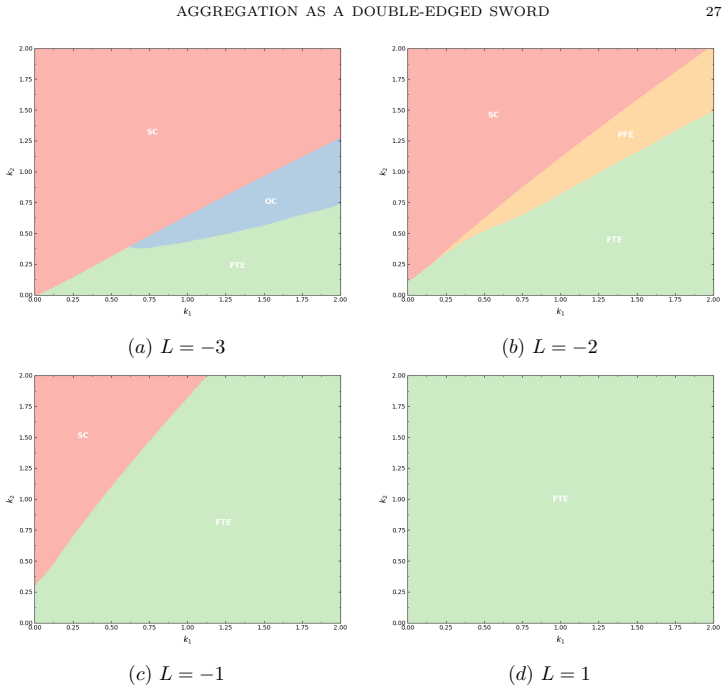
- Stronger aggregation enlarges the finite-time extinction region in the fear-Allee parameter plane.
- Weaker aggregation supports a richer set of coexistence outcomes including oscillatory dynamics.
- Finite-time extinction of the susceptible prey population triggers subsequent collapse of the infected prey and predator populations.
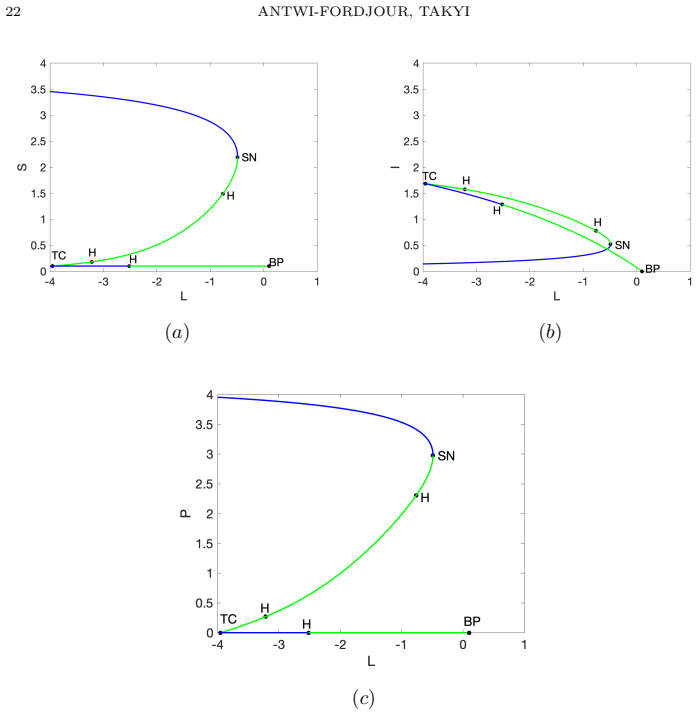
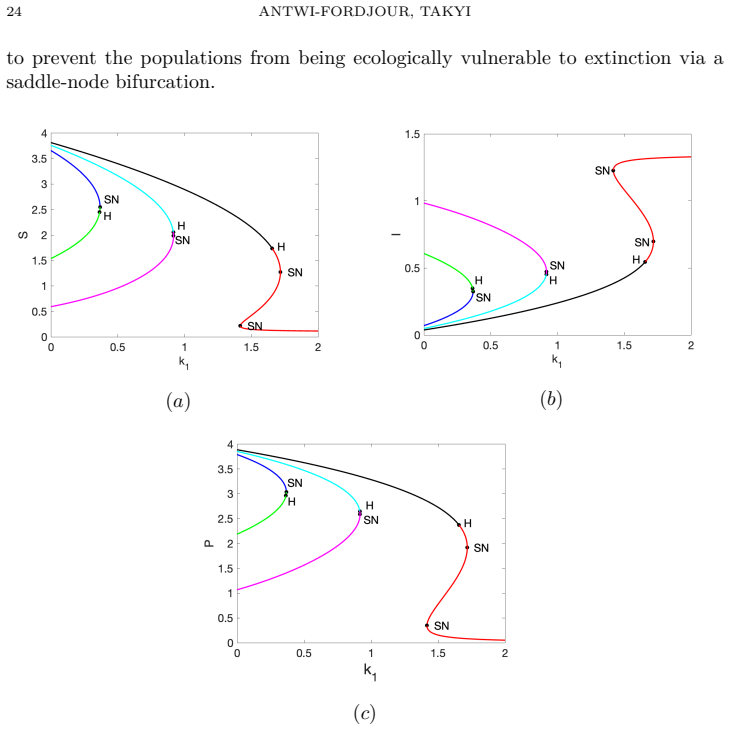
- Transcritical, saddle-node, and Hopf bifurcations occur as fear intensity, aggregation strength, and Allee threshold vary.
Where Pith is reading between the lines
- Field studies could test whether real extinction events occur faster in populations with high aggregation under disease pressure.
- Management interventions that alter grouping behavior might shift systems away from or toward the extinction region depending on disease presence.
- The bound suggests that parameter estimates from behavioral observations could predict collapse timelines without full model simulation.
Load-bearing premise
The sublinear aggregation-based predation term and the dual fear responses take the specific functional forms introduced in the model.
What would settle it
A direct measurement or simulation in which increasing aggregation strength lengthens rather than shortens the observed time to extinction, under matching fear and Allee conditions, would falsify the claimed upper bound.
Figures



read the original abstract
Prey aggregation is widely regarded as a defense against predation, yet we show that in disease-structured populations subject to predator-induced fear and demographic Allee thresholds, aggregation can paradoxically accelerate ecosystem collapse. We develop and analyze a susceptible-infectious-predator model incorporating dual fear responses -- together with a sublinear aggregation-based predation term and an Allee effect. Critically, we derive an explicit upper bound on the extinction time that decreases as predator pressure increases or aggregation strengthens, quantifying for the first time how behavioral and demographic parameters jointly determine the speed of ecological collapse. This finite-time extinction subsequently triggers a cascade collapse of the infected prey and predator populations, driving the entire ecological community to extinction. Bifurcation analysis reveals transcritical, saddle-node, and Hopf bifurcations as fear intensity, aggregation strength, and Allee threshold vary. Two-parameter continuation further identifies the precise regions of the fear--Allee parameter plane in which stable coexistence, oscillatory coexistence, predator exclusion, and finite-time extinction occur, demonstrating that stronger aggregation monotonically enlarges the finite-time extinction region while weaker aggregation supports a richer landscape of coexistence dynamics. These results demonstrate that behavioral defenses operating at the population level can generate abrupt ecological tipping points when they interact with disease dynamics and demographic vulnerability.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops a susceptible-infectious-predator ODE model that incorporates dual predator-induced fear responses, a sublinear aggregation-based predation term, and a demographic Allee threshold. It derives an explicit upper bound on the time to extinction of the susceptible prey population that decreases monotonically with predator pressure and aggregation strength; this finite-time extinction is shown to trigger cascade extinction of the infected prey and predator. Bifurcation analysis identifies transcritical, saddle-node, and Hopf bifurcations, while two-parameter continuation maps the fear–Allee plane into regions of stable coexistence, oscillatory coexistence, predator exclusion, and finite-time extinction, with the extinction region enlarging as aggregation strength increases.
Significance. If the explicit upper bound and its monotonicity hold under the stated assumptions, the work supplies the first quantitative estimate of how aggregation and fear jointly accelerate collapse in a disease-structured community, together with a complete bifurcation diagram of the tipping-point boundaries. The combination of an analytic extinction-time bound with numerical continuation of multiple bifurcation curves provides a concrete, parameter-dependent prediction that could be tested against field data on aggregation behavior and disease prevalence.
major comments (2)
- [model development] Model development section: the explicit upper bound on extinction time is obtained by constructing and integrating a differential inequality that relies on the precise sublinear form chosen for the aggregation predation term and the specific dual fear response functions; the paper provides no comparison with alternative biologically plausible forms (e.g., saturating or mildly superlinear aggregation), so the claimed monotonic decrease of the bound with aggregation strength is tied to these modeling choices rather than shown to be robust.
- [bifurcation analysis] Bifurcation analysis and two-parameter continuation: the statement that stronger aggregation monotonically enlarges the finite-time extinction region rests on the numerical continuation results, yet the manuscript supplies neither the continuation algorithm details, step-size tolerances, nor verification that the detected boundary curves are free of missed codimension-2 points; without these, the quantitative enlargement claim cannot be assessed for numerical reliability.
minor comments (1)
- The abstract asserts that the bound 'quantifies for the first time' the joint dependence; a brief literature pointer to prior analytic extinction-time estimates in fear or Allee models would strengthen this claim.
Simulated Author's Rebuttal
We thank the referee for their thorough review and constructive suggestions. We address each of the major comments below and will revise the manuscript accordingly where appropriate.
read point-by-point responses
-
Referee: [model development] Model development section: the explicit upper bound on extinction time is obtained by constructing and integrating a differential inequality that relies on the precise sublinear form chosen for the aggregation predation term and the specific dual fear response functions; the paper provides no comparison with alternative biologically plausible forms (e.g., saturating or mildly superlinear aggregation), so the claimed monotonic decrease of the bound with aggregation strength is tied to these modeling choices rather than shown to be robust.
Authors: The sublinear aggregation predation term is selected to reflect the biological reality that aggregation can reduce the per capita predation rate due to predator confusion or dilution effects, consistent with empirical observations in some prey species. The dual fear functions are derived from standard modeling of anti-predator behaviors affecting reproduction and survival. While the explicit bound and its monotonicity are rigorously derived for these forms, we acknowledge the value of assessing robustness. In the revised manuscript, we will add a paragraph in the discussion section addressing the choice of functional forms and noting that the qualitative conclusions may depend on these assumptions, without performing exhaustive comparisons which would extend the scope significantly. revision: partial
-
Referee: [bifurcation analysis] Bifurcation analysis and two-parameter continuation: the statement that stronger aggregation monotonically enlarges the finite-time extinction region rests on the numerical continuation results, yet the manuscript supplies neither the continuation algorithm details, step-size tolerances, nor verification that the detected boundary curves are free of missed codimension-2 points; without these, the quantitative enlargement claim cannot be assessed for numerical reliability.
Authors: The two-parameter continuations were carried out using standard numerical continuation software. To enhance reproducibility and address concerns about numerical reliability, we will include in the revised manuscript (or supplementary material) the specific algorithm employed, the step-size tolerances used, and a statement confirming that no additional codimension-2 bifurcation points were detected along the boundary curves within the explored parameter ranges. revision: yes
Circularity Check
No circularity: upper bound derived mathematically from explicitly stated model equations
full rationale
The paper defines a specific ODE model with chosen functional forms (sublinear aggregation predation, dual fear responses, Allee effect) in the model-development section, then applies standard comparison theorems and differential inequalities to those equations to obtain an explicit upper bound on extinction time. This is a direct consequence of integrating the constructed inequality; the bound is not equivalent to the inputs by definition, nor obtained by fitting parameters to data, nor justified solely by self-citation. The result quantifies behavior under the stated assumptions rather than smuggling an ansatz or renaming a prior result. The derivation chain is therefore self-contained against the model's own dynamics.
Axiom & Free-Parameter Ledger
free parameters (3)
- fear intensity parameters
- aggregation strength
- Allee threshold
axioms (2)
- domain assumption The chosen functional forms for dual fear responses and sublinear aggregation predation are biologically plausible.
- standard math Solutions of the ODE system exist globally until the extinction time.
Reference graph
Works this paper leans on
-
[1]
Ripple and Robert L
William J. Ripple and Robert L. Beschta. Wolves and the ecology of fear: Can predation risk structure ecosystems? BioScience, 54(8):755–766, 2004
2004
-
[2]
Beschta and William J
Robert L. Beschta and William J. Ripple. Return of wolves to yellowstone national park: Restoring a functionally intact ecological system. Restoration Ecology, 20(1):106–113, 2012
2012
-
[3]
Lima and Lawrence M
Steven L. Lima and Lawrence M. Dill. Behavioral decisions made under the risk of predation: A review and prospectus. Canadian Journal of Zoology , 68(4):619–640, 1990
1990
-
[4]
Relationships between direct predation and risk effects
Scott Creel and David Christianson. Relationships between direct predation and risk effects. Trends in Ecology & Evolution , 23(4):194–201, 2008
2008
-
[5]
Brown, John W
Joel S. Brown, John W. Laundré, and Mahesh Gurung. The ecology of fear: Optimal foraging, game theory, and trophic interactions. Journal of Mammalogy , 80(2):385–399, 1999
1999
-
[6]
Peacor and Earl E
Scott D. Peacor and Earl E. Werner. How dependent are species-pair interaction strengths on other species in the food web? Ecology, 85(10):2754–2765, 2004
2004
-
[7]
Zanette, Aija F
Liana Y. Zanette, Aija F. White, Marek C. Allen, and Michael Clinchy. Perceived predation risk reduces the number of offspring songbirds produce per year. Science, 334(6061):1398– 1401, 2011
2011
-
[8]
Zanette, and Xiaohua Zou
Xiaoying Wang, Liana Y. Zanette, and Xiaohua Zou. Modelling the fear effect in predator– prey interactions. Journal of Mathematical Biology , 73(5):1179–1204, 2016
2016
-
[9]
Population dynamics with multiple allee effects induced by fear fac- tors: A mathematical study on prey–predator interactions
Sourav Kumar Sasmal. Population dynamics with multiple allee effects induced by fear fac- tors: A mathematical study on prey–predator interactions. Applied Mathematical Modelling , 64:1–14, 2018. AGGREGATION AS A DOUBLE-EDGED SWORD 35
2018
-
[10]
Controlling chaos in three species food chain model with fear effect
Vikas Kumar and Nitu Kumari. Controlling chaos in three species food chain model with fear effect. AIMS Mathematics , 5(2):828–842, 2020
2020
-
[11]
Westmoreland, and Kendall H
Kwadwo Antwi-Fordjour, Sarah P. Westmoreland, and Kendall H. Bearden. Dual fear phe- nomenon in an eco-epidemiological model with prey aggregation. The European Physical Journal Plus , 139(6):518, 2024
2024
-
[12]
Anderson and Robert M
Roy M. Anderson and Robert M. May. Regulation and stability of host–parasite population interactions. i. regulatory processes. Journal of Animal Ecology , 47(1):219–247, 1978
1978
-
[13]
Hethcote
Herbert W. Hethcote. The mathematics of infectious diseases. SIAM Review , 42(4):599–653, 2000
2000
-
[14]
Chattopadhyay and O
J. Chattopadhyay and O. Arino. A predator–prey model with disease in the prey. Nonlinear Analysis: Theory, Methods & Applications , 36:747–766, 1999
1999
-
[15]
Epidemics in predator–prey models: Disease in the predators
Ezio Venturino. Epidemics in predator–prey models: Disease in the predators. IMA Journal of Mathematics Applied in Medicine and Biology , 19(3):185–205, 2002
2002
-
[16]
W. D. Hamilton. Geometry for the selfish herd. Journal of Theoretical Biology, 31(2):295–311, 1971
1971
-
[17]
Jens Krause and Graeme D. Ruxton. Living in Groups . Oxford University Press, Oxford, 2002
2002
-
[18]
Fear-driven extinction and (de) stabilization in a predator-prey model incorporating prey herd behavior and mutual interference
Kwadwo Antwi-Fordjour, Rana D Parshad, Hannah E Thompson, and Stephanie B Westaway. Fear-driven extinction and (de) stabilization in a predator-prey model incorporating prey herd behavior and mutual interference. AIMS Mathematics , 8(2):3353–3377, 2023
2023
-
[19]
P. A. Braza. Predator–prey dynamics with square root functional responses. Nonlinear Anal- ysis: Real World Applications , 13(4):1837–1843, 2012
2012
-
[20]
Prey herd behav- ior modeled by a generic non-differentiable functional response
Karina Vilches, Eduardo González-Olivares, and Alejandro Rojas-Palma. Prey herd behav- ior modeled by a generic non-differentiable functional response. Mathematical Modelling of Natural Phenomena , 13(3):26, 2018
2018
-
[21]
Animal Aggregations: A Study in General Sociology
Warder Clyde Allee. Animal Aggregations: A Study in General Sociology . University of Chicago Press, Chicago, 1931
1931
-
[22]
Inverse density dependence and the allee effect
Franck Courchamp, Tim Clutton-Brock, and Bryan Grenfell. Inverse density dependence and the allee effect. Trends in Ecology & Evolution , 14(10):405–410, 1999
1999
-
[23]
Allee Effects in Ecology and Con- servation
Franck Courchamp, Ludek Berec, and Joanna Gascoigne. Allee Effects in Ecology and Con- servation. Oxford University Press, Oxford, 2008
2008
-
[24]
Allee effects: Population growth, critical density, and the chance of extinction
Brian Dennis. Allee effects: Population growth, critical density, and the chance of extinction. Natural Resource Modeling, 3(4):481–538, 1989
1989
-
[25]
Stephens and William J
Philip A. Stephens and William J. Sutherland. Consequences of the allee effect for behaviour, ecology and conservation. Trends in Ecology & Evolution , 14(10):401–405, 1999
1999
-
[26]
Foley, Carl Folke, and Brian Walker
Marten Scheffer, Stephen Carpenter, Jonathan A. Foley, Carl Folke, and Brian Walker. Cat- astrophic shifts in ecosystems. Nature, 413:591–596, 2001
2001
-
[27]
Critical Transitions in Nature and Society
Marten Scheffer. Critical Transitions in Nature and Society . Princeton University Press, Princeton, 2009
2009
-
[28]
Hendry, Jonathan Levine, Nicolas Loeuille, Jon Norberg, Patrik Nosil, Marten Scheffer, and Luc De Meester
Vasilis Dakos, Blake Matthews, Andrew P. Hendry, Jonathan Levine, Nicolas Loeuille, Jon Norberg, Patrik Nosil, Marten Scheffer, and Luc De Meester. Ecosystem tipping points in an evolving world. Nature Ecology & Evolution , 3:355–362, 2019
2019
-
[29]
Dynamics of diseased-impacted prey populations: defense and allee effect mechanisms
Kwadwo Antwi-Fordjour, Zachary Overton, and Dylan Lee. Dynamics of diseased-impacted prey populations: defense and allee effect mechanisms. The European Physical Journal Plus , 140(7):675, 2025
2025
-
[30]
Differential equations and dynamical systems , volume 7
Lawrence Perko. Differential equations and dynamical systems , volume 7. Springer Science & Business Media, 2013
2013
-
[31]
Kuznetsov
Annick Dhooge, Willy Govaerts, and Yuri A. Kuznetsov. MATCONT: A MATLAB package for numerical bifurcation analysis of ODEs. ACM Transactions on Mathematical Software , 29(2):141–164, 2003
2003
-
[32]
Kuznetsov
Yuri A. Kuznetsov. Elements of Applied Bifurcation Theory . Springer, New York, 3 edition, 2004
2004
-
[33]
Brook, Navjot S
Barry W. Brook, Navjot S. Sodhi, and Corey J. A. Bradshaw. Synergies among extinction drivers under global change. Trends in Ecology & Evolution , 23(8):453–460, 2008
2008
-
[34]
Boerlijst, Tim Oudman, and André M
Melinda C. Boerlijst, Tim Oudman, and André M. de Roos. Catastrophic collapse can occur without early warning: Examples of silent catastrophes in structured ecological models. PLoS ONE, 8(4):e62033, 2013. 36 ANTWI-FORDJOUR, TAKYI
2013
-
[35]
Carpenter, William A
Vasilis Dakos, Stephen R. Carpenter, William A. Brock, Aaron M. Ellison, Vishwesha Guttal, Anthony R. Ives, Sonia Kéfi, Valentina Livina, David A. Seekell, Egbert H. van Nes, and Marten Scheffer. Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PLoS ONE , 7(7):e41010, 2012
2012
-
[36]
Some demographic and genetic consequences of environmental heterogeneity for biological control
Richard Levins. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bulletin of the Entomological Society of America , 15(3):237–240, 1969
1969
-
[37]
Metapopulation dynamics
Ilkka Hanski. Metapopulation dynamics. Nature, 396:41–49, 1998
1998
-
[38]
Risks of population extinction from demographic and environmental stochas- ticity and random catastrophes
Russell Lande. Risks of population extinction from demographic and environmental stochas- ticity and random catastrophes. The American Naturalist , 142(6):911–927, 1993
1993
-
[39]
Melbourne and Alan Hastings
Brett A. Melbourne and Alan Hastings. Extinction risk depends strongly on factors contribut- ing to stochasticity. Nature, 454:100–103, 2008
2008
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.