Synchronization modes in bipartite oscillator networks
Pith reviewed 2026-06-26 14:48 UTC · model grok-4.3
The pith
In a bipartite Kuramoto-Sakaguchi network, partial synchrony produces self-organized quasiperiodicity where one population decouples from the global field.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
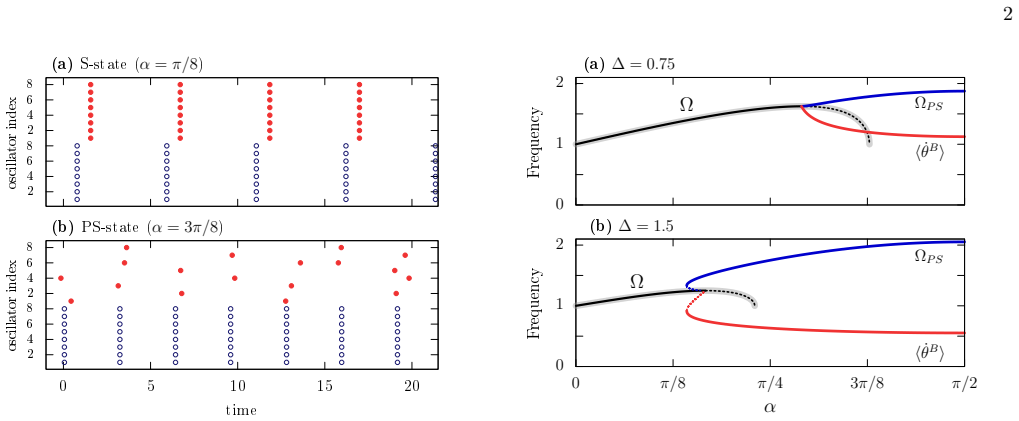
The partial synchrony state in the bipartite Kuramoto-Sakaguchi model constitutes self-organized quasiperiodicity. Global oscillations do not entrain one of the two populations, whose oscillators display quasiperiodic dynamics with an average frequency that can deviate significantly from the global field. This behavior arises in the canonical model despite its purely linear global coupling.
What carries the argument
The partial synchrony regime on the bipartite network, in which one population decouples into quasiperiodic motion while the other remains entrained to the linear global field.
If this is right
- Continuous and discontinuous transitions separate full synchrony from partial synchrony.
- One population exhibits quasiperiodic motion whose mean frequency deviates from the global oscillation.
- The decoupled population's frequency shift can be large while the other population remains phase-locked.
- Linear global coupling alone is sufficient to produce the quasiperiodic state without additional nonlinear terms.
Where Pith is reading between the lines
- Similar frequency mismatches could appear in any two-group oscillator system once a partial synchrony threshold is crossed.
- The mechanism offers a minimal explanation for observed rhythm differences between excitatory and inhibitory groups in neural data.
- Adding heterogeneity within each population might widen the parameter region where quasiperiodicity persists.
- The same bipartite structure could be tested in non-neural systems such as coupled chemical reactors or mechanical pendula.
Load-bearing premise
The bipartite Kuramoto-Sakaguchi model captures the coexistence of strongly and partially synchronized regimes observed in neuronal networks with excitatory and inhibitory populations.
What would settle it
A direct numerical integration of the bipartite equations that shows both populations always lock to the same average frequency in the partial synchrony regime would falsify the claim of self-organized quasiperiodicity.
Figures

read the original abstract
Collective oscillations in neuronal systems often arise from interactions between excitatory and inhibitory populations rather than from recurrent coupling within a single ensemble. Motivated by the coexistence of strongly and partially synchronized regimes in such systems, we study the Kuramoto Sakaguchi model on a bipartite network. Despite its minimal structure, the model exhibits rich collective dynamics, including both continuous and discontinuous transitions from full synchrony to partial synchrony (PS). In the PS regime, global oscillations fail to entrain one of the two populations, whose oscillators display quasiperiodic dynamics with an average frequency that can significantly deviate from that of the global field, as observed in neuronal networks. We show that this PS state constitutes an example of self-organized quasiperiodicity, arising here in the canonical Kuramoto Sakaguchi model despite its purely linear global coupling.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript studies the Kuramoto-Sakaguchi model on a bipartite network of two oscillator populations. It reports continuous and discontinuous transitions from full synchrony to a partial synchrony (PS) regime in which one population fails to entrain, producing quasiperiodic motion whose mean frequency deviates from that of the global field. The authors interpret the PS state as an instance of self-organized quasiperiodicity that appears in the canonical model despite its purely linear mean-field coupling.
Significance. If the reported transitions and frequency-offset quasiperiodicity are robust, the work supplies a minimal, analytically tractable example of how bipartite excitatory-inhibitory architecture can generate the coexistence of strongly and partially synchronized states observed in neuronal data. The demonstration that such behavior arises without additional nonlinearities or heterogeneous coupling strengths is a useful addition to the synchronization literature.
minor comments (3)
- [Abstract] Abstract and §1: the phrase 'self-organized quasiperiodicity' is introduced without an explicit definition or reference to prior usage; a one-sentence clarification of what distinguishes this usage from ordinary non-entrainment under linear drive would aid readers.
- [§3] §3 (or wherever the order-parameter equations appear): the global coupling is described as 'purely linear,' yet the Sakaguchi phase lag introduces a nonlinear sine term; a brief remark distinguishing the linearity of the mean-field interaction from the nonlinearity of the phase interaction would prevent misreading.
- [Figure 2] Figure 2 (or the panel showing the discontinuous transition): the caption should state the precise value of the phase-lag parameter α and the integration time step used to distinguish a true jump from a steep but continuous crossover.
Simulated Author's Rebuttal
We thank the referee for their positive summary and significance assessment of our work on the Kuramoto-Sakaguchi model on bipartite networks. We appreciate the recommendation of minor revision. No specific major comments appear in the report, so we have no point-by-point items to address.
Circularity Check
No significant circularity
full rationale
The paper is a forward simulation study of the standard bipartite Kuramoto-Sakaguchi model. Its central claim—that a partially synchronized state exhibits self-organized quasiperiodicity—is obtained by direct numerical integration of the model's equations under linear mean-field coupling. No parameters are fitted to the target dynamics, no predictions are made from subsets of the same data, and no load-bearing uniqueness theorems or ansatzes are imported via self-citation. The derivation chain therefore remains independent of its inputs and is self-contained.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
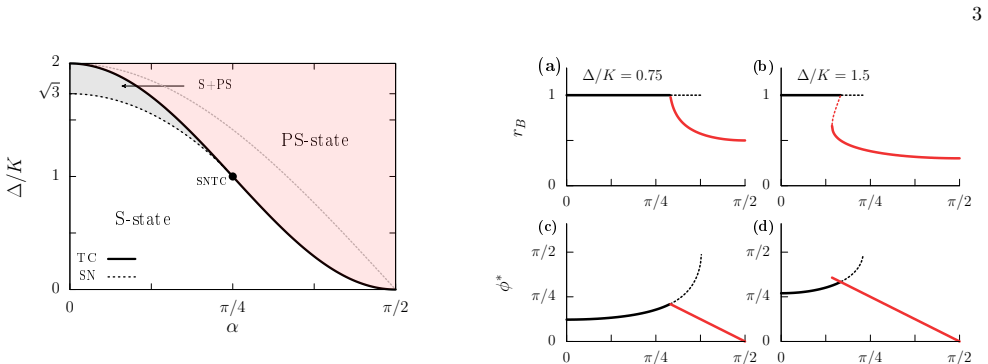
(5) is the locus of a transcritical (TC) bifurcation where the S and PS states exchange stability
Hence, Eq. (5) is the locus of a transcritical (TC) bifurcation where the S and PS states exchange stability
-
[2]
(14) shows that PS states are born withr B =K/∆≤1 in a SN bifurcation at αP S = arccos 1 2 p 1 + (∆/K)2 ,(15) for ∆/K∈(1, √ 3)
In addition, Eq. (14) shows that PS states are born withr B =K/∆≤1 in a SN bifurcation at αP S = arccos 1 2 p 1 + (∆/K)2 ,(15) for ∆/K∈(1, √ 3). The SN and TC bifurcations meet at a Saddle-Node-Transcritical (SNTC) bifurcation point whenr B =K/∆ = 1, forα=π/4. The unfolding of this codimension-two point reveals two scenarios — see Fig. 3. Specifically, th...
2021
-
[3]
Rosenblum and A
M. Rosenblum and A. Pikovsky, Self-organized quasiperi- odicity in oscillator ensembles with global nonlinear cou- pling, Phys. Rev. Lett.98, 064101 (2007)
2007
-
[4]
A. T. Winfree,The Geometry of Biological Time (Springer, New York, 1980)
1980
-
[5]
A. S. Pikovsky, M. G. Rosenblum, and J. Kurths,Syn- chronization, a Universal Concept in Nonlinear Sciences (Cambridge University Press, Cambridge, 2001)
2001
-
[6]
S. H. Strogatz,Sync: The emerging science of sponta- neous order.(Hyperion Press, New York, 2003)
2003
-
[7]
A. T. Winfree, Biological rhythms and the behavior of populations of coupled oscillators., J. Theor. Biol.16, 15 (1967)
1967
-
[8]
Kuramoto,Chemical Oscillations, Waves, and Turbu- lence(Springer-Verlag, Berlin, 1984)
Y. Kuramoto,Chemical Oscillations, Waves, and Turbu- lence(Springer-Verlag, Berlin, 1984)
1984
-
[9]
Fruchart, R
M. Fruchart, R. Hanai, P. B. Littlewood, and V. Vitelli, Non-reciprocal phase transitions, Nature592, 363 (2021). 5
2021
-
[10]
Whittington, R
M. Whittington, R. Traub, N. Kopell, B. Ermentrout, and E. Buhl, Inhibition-based rhythms: experimental and mathematical observations on network dynamics, Int. Journal of Psychophysiol.38, 315 (2000)
2000
-
[11]
B¨ orgers,An introduction to modeling neuronal dynam- ics, Vol
C. B¨ orgers,An introduction to modeling neuronal dynam- ics, Vol. 66 (Springer, 2017)
2017
-
[12]
Hansel and G
D. Hansel and G. Mato, Asynchronous states and the emergence of synchrony in large networks of interacting excitatory and inhibitory neurons, Neural Computation 15, 1 (2003)
2003
-
[13]
B¨ orgers and N
C. B¨ orgers and N. Kopell, Synchronization in Networks of Excitatory and Inhibitory Neurons with Sparse, Random Connectivity, Neural Computation15, 509 (2003)
2003
-
[14]
Montbri´ o and D
E. Montbri´ o and D. Paz´ o, Kuramoto model for excitation-inhibition-based oscillations, Phys. Rev. Lett. 120, 244101 (2018)
2018
-
[15]
Al- ternative modeling approaches have shown that PS states arise in E-I networks with random connectivity, noise, and synaptic delays [42, 43]
Previous studies have reproduced PS states in E-I net- works of spiking neurons by introducing stochastic inputs or additional currents in the E population [9, 44–48]. Al- ternative modeling approaches have shown that PS states arise in E-I networks with random connectivity, noise, and synaptic delays [42, 43]
-
[16]
Sakaguchi and Y
H. Sakaguchi and Y. Kuramoto, A soluble active rotator model showing phase transitions via mutual entrainment, Prog. Theor. Phys.76, 576 (1986)
1986
-
[17]
[12] a two-population Kuramoto model was de- rived from an E-I network withcurrent-basedsynapses, see also [23]
In Ref. [12] a two-population Kuramoto model was de- rived from an E-I network withcurrent-basedsynapses, see also [23]. This model without self-coupling (see foot- note [42] in Ref. [12]) corresponds to Eqs. (1) with αA,B = 0
-
[18]
Montbri´ o, J
E. Montbri´ o, J. Kurths, and B. Blasius, Synchronization of two interacting populations of oscillators, Phys. Rev. E70, 056125 (2004)
2004
-
[19]
D. M. Abrams, R. Mirollo, S. H. Strogatz, and D. A. Wiley, Phys. Rev. Lett.101, 084103 (2008)
2008
-
[20]
Pikovsky and M
A. Pikovsky and M. Rosenblum, Partially integrable dy- namics of hierarchical populations of coupled oscillators, Phys. Rev. Lett.101, 264103 (2008)
2008
-
[21]
C. R. Laing, Chimera states in heterogeneous networks, Chaos: An Interdisciplinary Journal of Nonlinear Science 19, 013113 (2009)
2009
-
[22]
M. J. Panaggio, D. M. Abrams, P. Ashwin, and C. R. Laing, Chimera states in networks of phase oscillators: The case of two small populations, Phys. Rev. E93, 012218 (2016)
2016
-
[23]
E. A. Martens, C. Bick, and M. J. Panaggio, Chimera states in two populations with heterogeneous phase-lag, Chaos: An Interdisciplinary Journal of Nonlinear Science 26, 094819 (2016)
2016
-
[24]
Kotwal, X
T. Kotwal, X. Jiang, and D. M. Abrams, Connecting the kuramoto model and the chimera state, Phys. Rev. Lett. 119, 264101 (2017)
2017
-
[25]
Clusella, B
P. Clusella, B. Pietras, and E. Montbri´ o, Kuramoto model for populations of quadratic integrate-and-fire neurons with chemical and electrical coupling, Chaos: An Interdisciplinary Journal of Nonlinear Science32, 013105 (2022)
2022
-
[26]
M. J. Panaggio and D. M. Abrams, Chimera states: Co- existence of coherence and incoherence in networks of coupled oscillators, Nonlinearity28, R67 (2015)
2015
-
[27]
Pikovsky and M
A. Pikovsky and M. Rosenblum, Dynamics of globally coupled oscillators: Progress and perspectives, Chaos25, 097616 (2015)
2015
-
[28]
C. Bick, M. Goodfellow, C. R. Laing, and E. A. Martens, Understanding the dynamics of biological and neural os- cillator networks through exact mean-field reductions: a review, The Journal of Mathematical Neuroscience10, 1 (2020)
2020
-
[29]
S. W. Haugland, The changing notion of chimera states, a critical review, J. Phys. Complex.2, 032001 (2021)
2021
-
[30]
Verwoerd and O
M. Verwoerd and O. Mason, On computing the critical coupling coefficient for the kuramoto model on a com- plete bipartite graph, SIAM Journal on Applied Dynam- ical Systems8, 417 (2009)
2009
-
[31]
Punetha, R
N. Punetha, R. Ramaswamy, and F. M. Atay, Bipartite networks of oscillators with distributed delays: Synchro- nization branches and multistability, Phys. Rev. E91, 042906 (2015)
2015
-
[32]
Chiba, G
H. Chiba, G. S. Medvedev, and M. S. Mizuhara, Bifur- cations in the kuramoto model on graphs, Chaos: An In- terdisciplinary Journal of Nonlinear Science28, 073109 (2018)
2018
-
[33]
U. S. Thounaojam, Explosive synchronization in bipar- tite networks, Chaos, Solitons and Fractals152, 111435 (2021)
2021
-
[34]
Y. Qin, M. Cao, B. D. O. Anderson, D. S. Bassett, and F. Pasqualetti, Mediated remote synchronization of kuramoto-sakaguchi oscillators: The number of me- diators matters, IEEE Control Systems Letters5, 767 (2021)
2021
-
[35]
Pikovsky and M
A. Pikovsky and M. Rosenblum, Self-organized partially synchronous dynamics in populations of nonlinearly cou- pled oscillators, Physica D: Nonlinear Phenomena238, 27 (2009)
2009
-
[36]
Le´ on and D
I. Le´ on and D. Paz´ o, Phase reduction beyond the first or- der: The case of the mean-field complex ginzburg-landau equation, Phys. Rev. E100, 012211 (2019)
2019
-
[37]
(1) becomeα A =α B =α, withα= (αA +α B)/2
Definingϑ B i =θ B i + (αA −α B)/2, the phase shift pa- rameters in Eq. (1) becomeα A =α B =α, withα= (αA +α B)/2. If the two population KS model has self- interactions,α A =α B cannot be adopted without loss of generality
-
[38]
Sakaguchi, S
H. Sakaguchi, S. Shinomoto, and Y. Kuramoto, Mutual Entrainment in Oscillator Lattices with Nonvariational Type Interaction, Progress of Theoretical Physics79, 1069 (1988)
1988
-
[39]
(1) gives one zero eigenvalue, and a (real) eigenvalue µ=d ˙ϕ/dϕ|ϕ=ϕ∗ =−K p (2 cosα)2 −(∆/K) 2; note that settingµ= 0 yields Eq
ForN= 1, the linear stability analysis of the S-state of Eqs. (1) gives one zero eigenvalue, and a (real) eigenvalue µ=d ˙ϕ/dϕ|ϕ=ϕ∗ =−K p (2 cosα)2 −(∆/K) 2; note that settingµ= 0 yields Eq. (2). ForN >1, there are two additional sets of (N−1)-times degenerated eigenvalues µA = 1 2(µ−∆ tanα) andµ B = 1 2(µ+ ∆ tanα), which describe the internal stability o...
-
[40]
Ott and T
E. Ott and T. M. Antonsen, Low dimensional behavior of large systems of globally coupled oscillators, Chaos18, 037113 (2008)
2008
-
[41]
Watanabe and S
S. Watanabe and S. H. Strogatz, Integrability of a glob- ally coupled oscillator array, Phys. Rev. Lett.70, 2391 (1993)
1993
-
[42]
(10-12) withr B = 1 andr A ≤1 exists in parameter regions not considered in the main text
An additional equilibrium of Eqs. (10-12) withr B = 1 andr A ≤1 exists in parameter regions not considered in the main text. See Supplemental Material for an analysis 6 valid for all ∆ and forα∈(−π/2, π/2)
-
[43]
The linear stability analysis of the PS-state of Eqs. (10-
-
[44]
The eigenvalues satisfy λ=λ + +λ − <0, and hence the only potential (station- ary) instability is throughλ +
yields three eigenvalues:λ=−Kr B sin(2α), and λ± =λ/2± q λ2/4 +K(K−r B∆) (1−r 2 B)/(2r 2 B), wherer B are given by Eq.(14). The eigenvalues satisfy λ=λ + +λ − <0, and hence the only potential (station- ary) instability is throughλ +. Note thatλ + = 0 when (i)r B = 1 (TC bif.) and when (ii)r B =K/∆ (SN bif.)
-
[45]
Brunel and V
N. Brunel and V. Hakim, Sparsely synchronized neuronal oscillations, Chaos: An Interdisciplinary Journal of Non- linear Science18, 015113 (2008)
2008
-
[46]
Wang, Neurophysiological and computational prin- ciples of cortical rhythms in cognition, Physiological Re- views90, 1195 (2010)
X.-J. Wang, Neurophysiological and computational prin- ciples of cortical rhythms in cognition, Physiological Re- views90, 1195 (2010)
2010
-
[47]
B¨ orgers, S
C. B¨ orgers, S. Epstein, and N. J. Kopell, Background gamma rhythmicity and attention in cortical local cir- cuits: A computational study, Proceedings of the Na- tional Academy of Sciences102, 7002 (2005)
2005
-
[48]
Kopell and G
N. Kopell and G. LeMasson, Rhythmogenesis, amplitude modulation, and multiplexing in a cortical architecture., Proceedings of the National Academy of Sciences91, 10586 (1994)
1994
-
[49]
Kilpatrick and B
Z. Kilpatrick and B. Ermentrout, Sparse gamma rhythms arising through clustering in adapting neuronal networks, PLoS Comput Biol7, e1002281 (2011)
2011
-
[50]
Krupa, S
M. Krupa, S. Gielen, and B. Gutkin, Adaptation and shunting inhibition leads to pyramidal/interneuron gamma with sparse firing of pyramidal cells, Journal of computational neuroscience37, 357 (2014)
2014
-
[51]
E. O. Mann, C. A. Radcliffe, and O. Paulsen, Hippocam- pal gamma-frequency oscillations: from interneurones to pyramidal cells, and back, The Journal of Physiology 562, 55 (2005)
2005
-
[52]
Lundqvist, A
M. Lundqvist, A. Compte, and A. Lansner, Bistable, ir- regular firing and population oscillations in a modular attractor memory network, PLOS Computational Biol- ogy6, 1 (2010)
2010
-
[53]
Dipoppa and B
M. Dipoppa and B. S. Gutkin, Flexible frequency control of cortical oscillations enables computations required for working memory, Proceedings of the National Academy of Sciences110, 12828 (2013)
2013
-
[54]
Dipoppa, M
M. Dipoppa, M. Szwed, and B. S. Gutkin, Controlling working memory operations by selective gating: the roles of oscillations and synchrony, Advances in cognitive psy- chology12, 209 (2016)
2016
-
[55]
Schmidt, D
H. Schmidt, D. Avitabile, E. Montbri´ o, and A. Roxin, Network mechanisms underlying the role of oscillations in cognitive tasks, PLoS Computational Biology14, 1 (2018)
2018
-
[56]
Y. G. Pavlov and B. Kotchoubey, Oscillatory brain activ- ity and maintenance of verbal and visual working mem- ory: A systematic review, Psychophysiology59, e13735 (2022)
2022
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.