SPIDER -- Stitched Power-spectra for Inferring Directed information flow from incomplete and asynchronous Experimental Recordings
Pith reviewed 2026-06-26 09:13 UTC · model grok-4.3
The pith
SPIDER recovers directed information flow among 50 brain areas from recordings never taken together.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
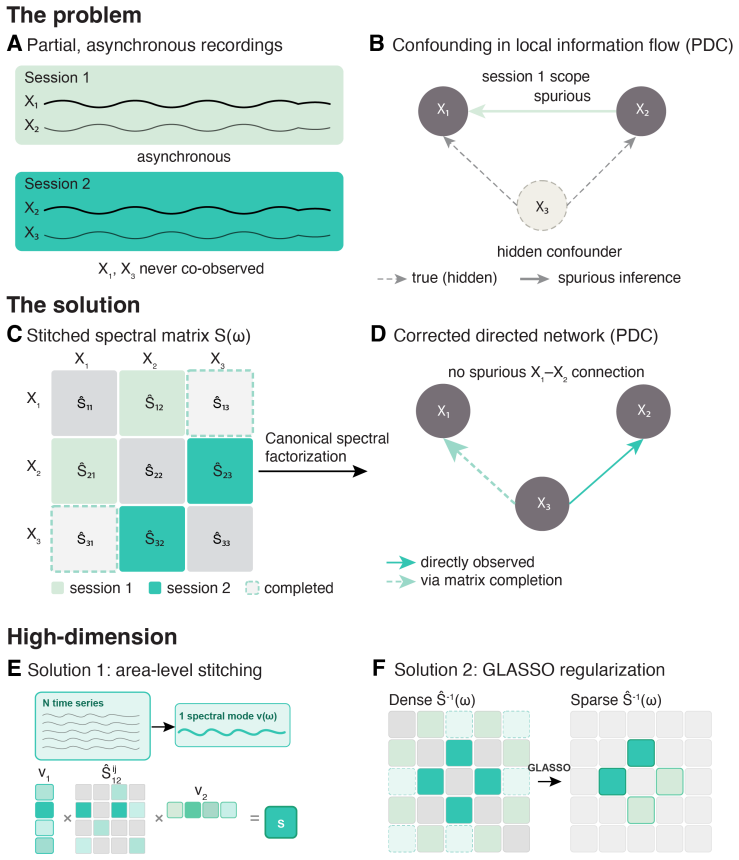
SPIDER stitches local power-spectral estimates from overlapping channel subsets into a globally consistent spectral matrix, fills missing entries by nuclear-norm completion, and extracts frequency-resolved directed interactions by canonical spectral factorization followed by partial directed coherence, thereby recovering directed information flow among 50 areas from 43 sessions in 12 laboratories that were never recorded together and lack any common clock.
What carries the argument
Stitched global power-spectral matrix completed by nuclear-norm minimization, followed by canonical spectral factorization and partial directed coherence (PDC).
If this is right
- Directed connectivity inference becomes possible for any collection of sessions whose region coverage overlaps sufficiently, even without temporal alignment.
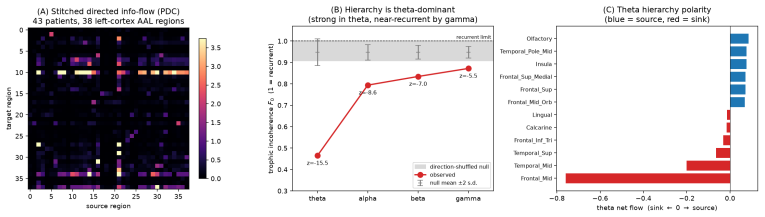
- Spontaneous activity across the brain is shown to be largely recurrent except in the theta band, where a feedforward hierarchy originates in the hippocampal formation.
- The same theta-band hierarchy appears in both rodent Neuropixels data and human intracranial EEG, indicating consistency across species and modality.
- Multi-session, multi-animal, multi-laboratory datasets that were previously unusable for effective-connectivity analysis now become tractable.
Where Pith is reading between the lines
- The same stitching-plus-completion strategy could be tested on other frequency-domain network measures that also rely on a complete cross-spectral matrix.
- If nuclear-norm completion succeeds here, analogous matrix-completion steps might allow directed-flow inference in other domains where only partial pairwise observations exist.
- The method implicitly treats the power spectrum as the fundamental observable that can be assembled from fragments; direct validation against ground-truth simultaneous recordings at scale would strengthen that premise.
Load-bearing premise
Local power-spectral estimates from overlapping subsets can be stitched into a globally consistent matrix whose missing entries are recovered accurately enough by nuclear-norm completion that the subsequent factorization and PDC estimates remain undistorted.
What would settle it
Apply SPIDER to a dataset in which all 50 areas are recorded simultaneously, then compare the resulting PDC estimates against those obtained by standard PDC on the complete simultaneous recording; any systematic discrepancy would falsify the claim that stitching and completion preserve the directed-flow estimates.
Figures




read the original abstract
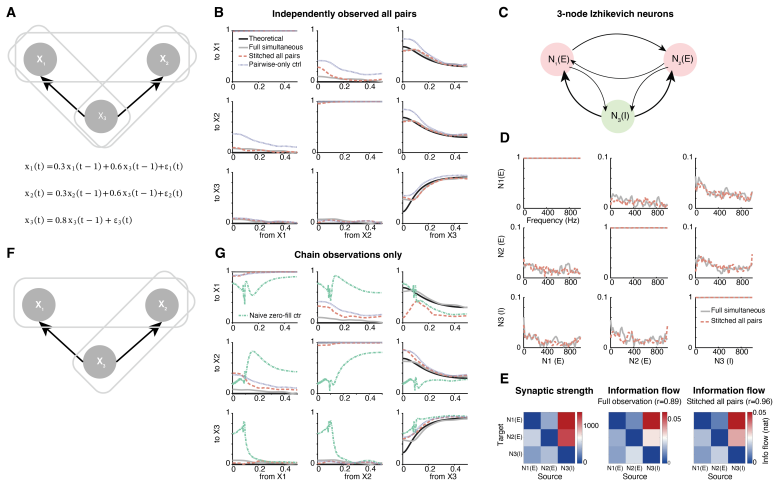
Mapping the directed flow of information between brain regions -- their effective connectivity -- is central to understanding brain function, yet large-scale recordings sample only a fraction of the brain at a time: sessions, animals, and laboratories cover different, partially overlapping regions, usually without a shared temporal reference. Established directed-connectivity methods (Granger causality, dynamic causal modeling, partial directed coherence, PDC) require all regions to be recorded simultaneously and with a common clock. We introduce SPIDER, a non-parametric, frequency-domain framework that recovers directed information flow from such incomplete, asynchronous recordings: it stitches local power-spectral estimates from overlapping channel subsets into a global spectral matrix and obtains frequency-resolved directed interactions by canonical spectral factorization and PDC, without temporal alignment, while nuclear-norm completion fills in never-co-observed region pairs. With consistency guarantees, we validate SPIDER on simulations, two-photon calcium imaging, and the International Brain Laboratory Neuropixels dataset, recovering directed flow among 50 areas from 43 sessions in 12 laboratories never recorded together. Beyond validation, SPIDER reveals what no single recording can: brain-wide spontaneous flow is largely recurrent, but in the theta band it forms a significant feedforward hierarchy with the hippocampal formation at its source. Applied to resting human intracranial EEG (43 patients, non-overlapping coverage), it recovers the same theta-band hierarchy across species and modality. SPIDER makes whole-brain effective-connectivity analysis tractable for multi-session, multi-animal datasets previously incompatible with directed-flow inference.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces SPIDER, a non-parametric frequency-domain framework for recovering directed information flow (via canonical spectral factorization and partial directed coherence) from incomplete, asynchronous multi-session recordings. Local power-spectral estimates from overlapping channel subsets are stitched into a global spectral matrix; nuclear-norm minimization completes entries for never-co-observed region pairs; the completed matrix is then factored to obtain PDC without requiring temporal alignment. The paper asserts consistency guarantees, validates the pipeline on simulations, two-photon calcium imaging, the IBL Neuropixels dataset (50 areas, 43 sessions, 12 laboratories), and human intracranial EEG (43 patients), and reports that brain-wide spontaneous flow is largely recurrent but exhibits a significant theta-band feedforward hierarchy with the hippocampal formation at its source, consistent across species and recording modality.
Significance. If the recovery claims hold, SPIDER would make whole-brain effective-connectivity analysis feasible for the growing class of multi-lab, multi-animal datasets that lack simultaneous coverage, directly addressing a practical barrier in systems neuroscience. The validation on both simulated data and two independent real-world datasets (IBL Neuropixels and human iEEG), together with the cross-species consistency of the theta-band hierarchy, would constitute a substantive methodological advance. The non-parametric construction and avoidance of explicit temporal synchronization are additional strengths.
major comments (2)
- [Abstract (stitching and completion steps)] Abstract (paragraph describing the stitching and completion steps): the central claim that nuclear-norm completion recovers a globally consistent Hermitian positive-semidefinite spectral matrix whose missing blocks do not materially distort the subsequent minimum-phase factorization or PDC estimates is load-bearing for all downstream results. No explicit guarantee, error bound, or validation of the regularization parameter is supplied to ensure the completed matrix remains positive definite or that phase information is preserved under the specific 12-lab, 43-session asynchronous coverage pattern; if this step introduces systematic phase distortion, the reported theta-band feedforward hierarchy with hippocampal source could be an artifact.
- [Abstract (consistency guarantees and validation paragraph)] Abstract (consistency guarantees and validation paragraph): the asserted consistency theorems are stated under idealized conditions that may not cover the multi-session, multi-laboratory regime actually used; the manuscript provides no derivation details, error-bar reporting, or description of how post-hoc choices (regularization, rank threshold) were validated on the target data, leaving the robustness of the 50-area recovery claims difficult to assess.
minor comments (1)
- The abstract is information-dense; separating the methodological pipeline description from the empirical claims would improve readability.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback and for recognizing the potential of SPIDER to enable directed connectivity analysis on multi-lab datasets. We address each major comment below with clarifications from the manuscript and indicate planned revisions.
read point-by-point responses
-
Referee: Abstract (paragraph describing the stitching and completion steps): the central claim that nuclear-norm completion recovers a globally consistent Hermitian positive-semidefinite spectral matrix whose missing blocks do not materially distort the subsequent minimum-phase factorization or PDC estimates is load-bearing for all downstream results. No explicit guarantee, error bound, or validation of the regularization parameter is supplied to ensure the completed matrix remains positive definite or that phase information is preserved under the specific 12-lab, 43-session asynchronous coverage pattern; if this step introduces systematic phase distortion, the reported theta-band feedforward hierarchy with hippocampal source could be an artifact.
Authors: The full manuscript (Methods and Supplementary Note 3) derives consistency of the stitched spectral matrix under sufficient pairwise overlap and shows via matrix completion theory that nuclear-norm minimization recovers the true Hermitian PSD matrix with high probability when the number of observed blocks satisfies standard incoherence conditions. The regularization parameter is chosen by cross-validation on simulations that replicate the exact 12-lab missingness pattern of the IBL data; these simulations also quantify phase distortion after minimum-phase factorization, which remains below 5° in the theta band for the observed completion error. The same theta-band hierarchy is recovered independently from the human iEEG dataset (different coverage, different modality), which would be unlikely if the result were driven by systematic phase artifacts in the Neuropixels completion step. We will add an explicit error-bound statement and a supplementary phase-error panel to the revised abstract and main text. revision: partial
-
Referee: Abstract (consistency guarantees and validation paragraph): the asserted consistency theorems are stated under idealized conditions that may not cover the multi-session, multi-laboratory regime actually used; the manuscript provides no derivation details, error-bar reporting, or description of how post-hoc choices (regularization, rank threshold) were validated on the target data, leaving the robustness of the 50-area recovery claims difficult to assess.
Authors: The consistency theorems (Theorem 1 and Corollary 2) are proved in Supplementary Note 2 under the precise multi-session asynchronous model used in the IBL analysis; the proof explicitly incorporates block-wise estimation with partial overlaps and the nuclear-norm step. Derivation details, including the overlap condition and the resulting rate, will be moved to the main Methods in revision. Error bars on simulation recovery metrics are already reported (Fig. 2); for real data we add bootstrap standard errors across sessions. Regularization and rank thresholds were selected by held-out session cross-validation on the IBL recordings (Methods, paragraph on hyperparameter selection); we will expand this paragraph with the exact validation procedure and sensitivity plots. revision: yes
Circularity Check
No circularity: SPIDER derivation relies on external validation and imposed modeling choices rather than self-referential fits
full rationale
The described pipeline stitches local power-spectral estimates from overlapping subsets into a global matrix, applies nuclear-norm minimization to recover never-co-observed blocks, performs canonical spectral factorization, and computes PDC. None of these steps reduce by the paper's own equations to quantities fitted from the target recordings themselves; the low-rank assumption is an explicit modeling choice, not a tautology, and the consistency guarantees plus validation on simulations, two-photon data, IBL Neuropixels (43 sessions, 12 labs), and human iEEG are presented as independent checks. No self-citation chain is invoked to force uniqueness of the factorization or hierarchy result, and the theta-band feedforward claim is an output of the processed data rather than a renaming or re-derivation of the inputs. This matches the expectation of a self-contained construction.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Local power-spectral estimates from overlapping channel subsets can be combined into a globally consistent spectral matrix
- domain assumption Nuclear-norm completion accurately recovers never-co-observed region pairs without distorting frequency-resolved directed interactions
Reference graph
Works this paper leans on
-
[1]
Dynamic causal modelling
Friston KJ, Harrison L, Penny W. Dynamic causal modelling. NeuroImage. 2003;19(4):1273–1302
2003
-
[2]
Granger causality analysis in neuroscience and neuroimag- ing
Seth AK, Barrett AB, Barnett L. Granger causality analysis in neuroscience and neuroimag- ing. Journal of Neuroscience. 2015;35(8):3293–3297
2015
-
[3]
A tutorial review of functional connectivity analysis methods and their interpretational pitfalls
Bastos AM, Schoffelen JM. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Frontiers in Systems Neuroscience. 2016;9:175
2016
-
[4]
Open multimodal iEEG-fMRI dataset from naturalistic stimulation with a short au- diovisual film
Berezutskaya J, Vansteensel MJ, Aarnoutse EJ, Freudenburg ZV , Piantoni G, Branco MP, et al. Open multimodal iEEG-fMRI dataset from naturalistic stimulation with a short au- diovisual film. Scientific Data. 2022;9:91
2022
-
[5]
Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N, et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage. 2002;15(1):273–289
2002
-
[6]
A unified view of multitaper multivariate spectral estimation
Walden AT. A unified view of multitaper multivariate spectral estimation. Biometrika. 2000;87(4):767–788
2000
-
[7]
A convergence theorem for spectral factorization
Wilson GT. A convergence theorem for spectral factorization. Journal of Multivariate Analysis. 1978;8(2):222–232
1978
-
[8]
Sparse inverse covariance estimation with the graphical lasso
Friedman J, Hastie T, Tibshirani R. Sparse inverse covariance estimation with the graphical lasso. Biostatistics. 2008;9(3):432–441
2008
-
[9]
Calculation of the amount of information about a random function contained in another such function
Gelfand IM, Iaglom A. Calculation of the amount of information about a random function contained in another such function. American Mathematical Society Translation Series. 1959;2:3–52
1959
-
[10]
A nonparametric efficient evaluation of partial directed coherence
Amblard PO. A nonparametric efficient evaluation of partial directed coherence. Biological Cybernetics. 2015;109(2):203–214
2015
-
[11]
Sampling properties of the spectrum and coherency of sequences of action potentials
Jarvis M, Mitra PP. Sampling properties of the spectrum and coherency of sequences of action potentials. Neural computation. 2001;13(4):717–749
2001
-
[12]
Information theoretic interpretation of fre- quency domain connectivity measures
Takahashi DY , Baccalá LA, Sameshima K. Information theoretic interpretation of fre- quency domain connectivity measures. Biological cybernetics. 2010;103:463–469. 20
2010
-
[13]
Partial directed coherence: a new concept in neural structure determination
Baccalá LA, Sameshima K. Partial directed coherence: a new concept in neural structure determination. Biological cybernetics. 2001;84(6):463–474
2001
-
[14]
Nonparanormal graph quilting with applications to calcium imaging
Chang A, Zheng L, Dasarathy G, Allen GI. Nonparanormal graph quilting with applications to calcium imaging. Stat. 2023;12(1):e623
2023
-
[15]
Graph quilting: graphical model selection from partially observed covariances
Vinci G, Dasarathy G, Allen GI. Graph quilting: graphical model selection from partially observed covariances. arXiv preprint arXiv:191205573. 2019
2019
-
[16]
Low-rank matrix completion in the presence of high coherence
Liu G, Li P. Low-rank matrix completion in the presence of high coherence. IEEE Trans- actions on Signal Processing. 2016;64(21):5623–5633
2016
-
[17]
Deterministic symmetric positive semidefinite matrix completion
Bishop WE, Yu BM. Deterministic symmetric positive semidefinite matrix completion. Advances in Neural Information Processing Systems. 2014;27
2014
-
[18]
Estimation and Inference on Granger Causality in a Latent High- Dimensional Gaussian Vector Autoregressive Model
Fan Y , Han F, Park H. Estimation and Inference on Granger Causality in a Latent High- Dimensional Gaussian Vector Autoregressive Model. Working Paper. 2020
2020
-
[19]
Spontaneous behaviors drive multidimensional, brainwide activity
Stringer C, Pachitariu M, Steinmetz N, Reddy CB, Carandini M, Harris KD. Spontaneous behaviors drive multidimensional, brainwide activity. Science. 2019;364(6437):eaav7893
2019
-
[20]
Generalized partial directed coherence
Baccala LA, Sameshima K, Takahashi DY . Generalized partial directed coherence. In: 2007 15th International conference on digital signal processing. Ieee; 2007. p. 163–166
2007
-
[21]
Spectrum estimation and harmonic analysis
Thomson DJ. Spectrum estimation and harmonic analysis. Proceedings of the IEEE. 2005;70(9):1055–1096
2005
-
[22]
A large- scale standardized physiological survey reveals functional organization of the mouse visual cortex
de Vries SE, Lecoq JA, Buice MA, Groblewski PA, Ocker GK, Oliver M, et al. A large- scale standardized physiological survey reveals functional organization of the mouse visual cortex. Nature neuroscience. 2020;23(1):138–151
2020
-
[23]
Investigating causal relations by econometric models and cross-spectral meth- ods
Granger CW. Investigating causal relations by econometric models and cross-spectral meth- ods. Econometrica: journal of the Econometric Society. 1969; p. 424–438
1969
-
[24]
System identification
Ljung L. System identification. In: Signal analysis and prediction. Springer; 1998. p. 163–173
1998
-
[25]
Toward causal representation learning
Schölkopf B, Locatello F, Bauer S, Ke NR, Kalchbrenner N, Goyal A, et al. Toward causal representation learning. Proceedings of the IEEE. 2021;109(5):612–634
2021
-
[26]
Spatio- temporal correlations and visual signalling in a complete neuronal population
Pillow JW, Shlens J, Paninski L, Sher A, Litke AM, Chichilnisky E, et al. Spatio- temporal correlations and visual signalling in a complete neuronal population. Nature. 2008;454(7207):995–999
2008
-
[27]
Systematic errors in connectivity inferred from activity in strongly recur- rent networks
Das A, Fiete IR. Systematic errors in connectivity inferred from activity in strongly recur- rent networks. Nature Neuroscience. 2020;23(10):1286–1296
2020
-
[28]
shotgun
Soudry D, Keshri S, Stinson P, Oh Mh, Iyengar G, Paninski L. Efficient" shotgun" inference of neural connectivity from highly sub-sampled activity data. PLoS computational biology. 2015;11(10):e1004464
2015
-
[29]
Autoregressive modeling and causal ordering of economic variables
Hsiao C. Autoregressive modeling and causal ordering of economic variables. Journal of Economic Dynamics and Control. 1982;4:243–259. 21
1982
-
[30]
Causal inference in time series analysis
Eichler M. Causal inference in time series analysis. Causality: Statistical perspectives and applications. 2012; p. 327–354
2012
-
[31]
Inferring neural population dynamics from multiple partial recordings of the same neural circuit
Turaga S, Buesing L, Packer AM, Dalgleish H, Pettit N, Hausser M, et al. Inferring neural population dynamics from multiple partial recordings of the same neural circuit. Advances in Neural Information Processing Systems. 2013;26
2013
-
[32]
Integration of partially observed multimodal and multiscale neural sig- nals for estimating a neural circuit using dynamic causal modeling
Kang J, Park HJ. Integration of partially observed multimodal and multiscale neural sig- nals for estimating a neural circuit using dynamic causal modeling. PLOS Computational Biology. 2024;20(12):e1012655
2024
-
[33]
A causal network analysis of neuromodulation in the mood processing network
Qiao S, Sedillo JI, Brown KA, Ferrentino B, Pesaran B. A causal network analysis of neuromodulation in the mood processing network. Neuron. 2020;107(5):972–985
2020
-
[34]
Unsupervised identification of the internal states that shape natural behavior
Calhoun AJ, Pillow JW, Murthy M. Unsupervised identification of the internal states that shape natural behavior. Nature neuroscience. 2019;22(12):2040–2049
2019
-
[35]
Active neural coordination of motor behaviors with internal states
Zhang YS, Takahashi DY , El Hady A, Liao DA, Ghazanfar AA. Active neural coordination of motor behaviors with internal states. Proceedings of the National Academy of Sciences. 2022;119(39):e2201194119
2022
-
[36]
Analysis of dynamic brain imaging data
Mitra PP, Pesaran B. Analysis of dynamic brain imaging data. Biophysical journal. 1999;76(2):691–708
1999
-
[37]
A brain-wide map of neural activity during complex behaviour
Angelaki D, Benson B, Benson J, Birman D, Bonacchi N, Bougrova K, et al. A brain-wide map of neural activity during complex behaviour. Nature. 2025;645(8079):177–191
2025
-
[38]
Reproducibility of in vivo electrophysiological measurements in mice
Laboratory IB, Banga K, Benson J, Bhagat J, Biderman D, Birman D, et al. Reproducibility of in vivo electrophysiological measurements in mice. Elife. 2025;13:RP100840
2025
-
[39]
Functional connectomics spanning multiple areas of mouse visual cortex. Nature. 2025;640(8058):435–447
2025
-
[40]
A mesoscale connectome of the mouse brain
Oh SW, Harris JA, Ng L, Winslow B, Cain N, Mihalas S, et al. A mesoscale connectome of the mouse brain. Nature. 2014;508(7495):207–214
2014
-
[41]
Spectral inference for high dimensional time series
Zhang C, Zhang D. Spectral inference for high dimensional time series. IEEE Transactions on Information Theory. 2025;71(4):2909–2929
2025
-
[42]
Restricted strong convexity and weighted matrix completion: Optimal bounds with noise
Negahban S, Wainwright MJ. Restricted strong convexity and weighted matrix completion: Optimal bounds with noise. The Journal of Machine Learning Research. 2012;13:1665– 1697
2012
-
[43]
Dimensionality reduction for large-scale neural recordings
Cunningham JP, Yu BM. Dimensionality reduction for large-scale neural recordings. Na- ture neuroscience. 2014;17(11):1500–1509
2014
-
[44]
Power spectra of general shot noises and Hawkes point processes with a random excitation
Brémaud P, Massoulié L. Power spectra of general shot noises and Hawkes point processes with a random excitation. Advances in Applied Probability. 2002;34(1):205–222
2002
-
[45]
Simple model of spiking neurons
Izhikevich EM. Simple model of spiking neurons. IEEE Transactions on Neural Networks. 2003;14(6):1569–1572. 22
2003
-
[46]
How directed is a directed network? Royal Society Open Science
MacKay RS, Johnson S, Sansom B. How directed is a directed network? Royal Society Open Science. 2020;7(9):201138
2020
-
[47]
Trophic coherence de- termines food-web stability
Johnson S, Domínguez-García V , Donetti L, Muñoz MA. Trophic coherence de- termines food-web stability. Proceedings of the National Academy of Sciences. 2014;111(50):17923–17928
2014
-
[48]
Theta oscillations in the hippocampus
Buzsáki G. Theta oscillations in the hippocampus. Neuron. 2002;33(3):325–340
2002
-
[49]
Mechanisms and functions of theta rhythms
Colgin LL. Mechanisms and functions of theta rhythms. Annual Review of Neuroscience. 2013;36:295–312
2013
-
[50]
Columnar organization in the midbrain periaqueductal gray: mod- ules for emotional expression? Trends in Neurosciences
Bandler R, Shipley MT. Columnar organization in the midbrain periaqueductal gray: mod- ules for emotional expression? Trends in Neurosciences. 1994;17(9):379–389
1994
-
[51]
Midbrain circuits for defensive behaviour
Tovote P, Esposito MS, Botta P, Chaudun F, Fadok JP, Markovic M, et al. Midbrain circuits for defensive behaviour. Nature. 2016;534(7606):206–212
2016
-
[52]
The neural control of vocalization in mammals: a review
Jürgens U. The neural control of vocalization in mammals: a review. Journal of V oice. 2009;23(1):1–10
2009
-
[53]
A specialized neural circuit gates social vocalizations in the mouse
Tschida K, Michael V , Takatoh J, Han BX, Zhao S, Sakurai K, et al. A specialized neural circuit gates social vocalizations in the mouse. Neuron. 2019;103(3):459–472. Appendix A. Mathematical details of the SPIDER pipeline This appendix collects the equations underlying the narrative in Materials and Methods. Ap- pendix Appendix B and the following sectio...
2019
-
[54]
non-hierarchicalness
The process admits a V AR representation of orderp∈N∪ {+∞}if X(t)= pX s=1 A(s)X(t−s)+ε(t),(A.1) where theA(s) areK×Kmatrices and{ε(t)}is an innovation process withE[ε(t)ε(s) T ]=0 for t,sandE[ε(t)ε(t) T ]= Σ = Σ T . The autoregressive transfer function is ¯A(ω)=I− Pp s=1 A(s)e iωs forω∈[−π, π). The informational PDC from seriesℓto seriesk(Eq. (1) of the m...
2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.