A Nonequilibrium Internal-Time Model of Aging: Entropy-Normalized Biological Proper Time and Repair Bifurcations
Pith reviewed 2026-06-26 06:03 UTC · model grok-4.3
The pith
Biological aging tracks the consumption of an internal entropy-cycle budget rather than elapsed calendar time.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The central claim is that aging is governed by the accumulation of internal physiological time rather than chronological time alone. The entropy-normalized extension is defined as Tσ(t) = ∫ [σz(s)/σref] f(s) ds, where f(s) is instantaneous physiological frequency and σz(s) is entropy produced per cycle. The normalized PBTE age is then APBTE(t) = Tσ(t)/Nref, which directly measures the fraction of a reference lifetime entropy-cycle budget that has been consumed.
What carries the argument
The entropy-normalized biological proper time Tσ(t), which integrates the product of physiological frequency and entropy cost per cycle, normalized to a reference unit.
If this is right
- Individuals with identical calendar age can be assigned different biological ages when their integrated entropy-weighted cycle counts differ.
- Species with different baseline physiological frequencies or entropy costs per cycle will have different mappings from chronological time to APBTE age.
- Repair capacity can be modeled as a function of remaining fraction of the reference entropy budget rather than of calendar time.
- Mortality hazard can be expressed as a function of APBTE age, allowing direct comparison across lifestyles that alter f(s) or σz(s).
Where Pith is reading between the lines
- The coordinate could be tested by checking whether interventions that lower average heart rate or metabolic rate slow the rise of APBTE relative to calendar time.
- If the model holds, biomarkers calibrated to Tσ(t) rather than to years lived might improve lifespan prediction in cohorts with high variance in activity or stress.
- The framework invites extension to non-mammalian systems where cycle counts such as heartbeats or breaths can still be recorded and entropy costs estimated.
Load-bearing premise
That physiological frequency and entropy produced per cycle can be defined and integrated to represent aging progress independently of chronological time.
What would settle it
Two individuals or species that reach identical values of Tσ(t) yet exhibit systematically different remaining lifespans, molecular damage accumulation, or mortality hazards.
Figures



read the original abstract
Chronological age is an incomplete coordinate for aging. Individuals and species sharing the same calendar time can differ substantially in physiological reserve, molecular damage, mortality hazard, and remaining lifespan. The Principle of Biological Time Equivalence (PBTE) offers a thermodynamic reformulation: biological aging is governed by the accumulation of \emph{internal} physiological time rather than chronological time alone. Building on prior PBTE work, this paper defines the internal-time coordinate $\theta(t)=\int_0^t f(s)\dd s$, where $t$ is chronological time and $f(s)$ is an instantaneous physiological frequency (for example heart rate or respiratory rate), so that $\theta$ is the accumulated count of physiological cycles. Its entropy-normalized extension is $\Tsig(t)=\int_0^t[\sigz(s)/\sref]f(s)\dd s$, where $\sigz(s)=\dd\Sigma/\dd\theta$ is the entropy produced per physiological cycle (the entropy cost per biological tick), $\Sigma$ is cumulative entropy production, and $\sref$ is a fixed reference entropy cost per cycle used as a normalizing unit. The normalized PBTE age $\APBTE(t)=\Tsig(t)/\Nref$ measures the fraction of a reference entropy--cycle budget consumed, where $\Nref$ is the reference number of entropy-weighted cycles available over a lifetime. The manuscript is explicitly theoretical: no empirical cohort is analyzed, and the numerical demonstrations are synthetic stress tests rather than validation.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes the Principle of Biological Time Equivalence (PBTE) as a thermodynamic reformulation of aging, defining an internal-time coordinate θ(t) = ∫ f(s) ds (accumulated physiological cycles), its entropy-normalized extension Tσ(t) = ∫ [σz(s)/sref] f(s) ds where σz(s) = dΣ/dθ is entropy cost per cycle, and the normalized age APBTE(t) = Tσ(t)/Nref as the fraction of a reference entropy-cycle budget consumed. The manuscript is explicitly theoretical, offering only definitional constructions and synthetic stress tests with no empirical cohort or validation against observables.
Significance. If the functions f(s) and σz(s) could be independently derived from measurable physiology (e.g., heart rate, metabolic rates) and first-principles nonequilibrium thermodynamics, the framework might supply a new internal coordinate for aging that decouples physiological progress from chronological time. The current manuscript, however, supplies no such derivation, mapping, or closure relation, so the significance remains limited to conceptual introduction.
major comments (2)
- [Abstract] Abstract: The central claim that Tσ(t) and APBTE(t) govern aging independently of chronological t requires that f(s) and σz(s) be obtainable from observables without circular appeal to aging outcomes; no such mapping, thermodynamic derivation, or closure relation is supplied, leaving the coordinate equivalent to the input definitions by construction.
- [Abstract] Abstract and synthetic stress tests: APBTE(t) is defined as Tσ(t)/Nref with fixed reference values sref and Nref chosen to normalize the scale; this renders the reported 'age' measure tautological with the chosen inputs, providing no external benchmark or falsifiable prediction.
Simulated Author's Rebuttal
We thank the referee for the careful reading of our manuscript on the Principle of Biological Time Equivalence (PBTE). The comments correctly identify the theoretical character of the work. We respond to each major comment below.
read point-by-point responses
-
Referee: [Abstract] Abstract: The central claim that Tσ(t) and APBTE(t) govern aging independently of chronological t requires that f(s) and σz(s) be obtainable from observables without circular appeal to aging outcomes; no such mapping, thermodynamic derivation, or closure relation is supplied, leaving the coordinate equivalent to the input definitions by construction.
Authors: The manuscript is explicitly a theoretical introduction of definitional constructions for the internal coordinates θ(t), Tσ(t), and APBTE(t). The claimed independence from chronological time follows directly from the integral forms, which accumulate according to the physiological frequency f(s) and entropy cost per cycle σz(s) rather than t. We agree that no empirical mapping, first-principles derivation, or closure relation is supplied; the paper states in the abstract and text that it contains only definitional constructions and synthetic stress tests. The contribution is the formal thermodynamic reformulation itself. No revision is required. revision: no
-
Referee: [Abstract] Abstract and synthetic stress tests: APBTE(t) is defined as Tσ(t)/Nref with fixed reference values sref and Nref chosen to normalize the scale; this renders the reported 'age' measure tautological with the chosen inputs, providing no external benchmark or falsifiable prediction.
Authors: The fixed reference values sref and Nref are introduced deliberately to produce the normalized, dimensionless quantity APBTE(t) as the fraction of a reference entropy-cycle budget. This normalization is part of the definition and is standard for constructing relative age measures. The synthetic stress tests are presented only to illustrate qualitative behaviors of the defined quantities under varying f(s) and σz(s), not as empirical validation or falsifiable predictions. The manuscript makes no claim to external benchmarks or predictions, consistent with its stated theoretical scope. No revision is required. revision: no
Circularity Check
APBTE(t) age measure defined by construction as normalized entropy-cycle integral
specific steps
-
self definitional
[Abstract]
"The normalized PBTE age APBTE(t)=Tsig(t)/Nref measures the fraction of a reference entropy--cycle budget consumed, where Nref is the reference number of entropy-weighted cycles available over a lifetime. ... Its entropy-normalized extension is Tsig(t)=∫[σz(s)/σref]f(s)ds, where σz(s)=dΣ/dθ ..."
APBTE(t) is introduced as the quantity that 'measures the fraction' but is simultaneously defined to be exactly that normalized integral; the reported age coordinate is therefore identical to its own input definitions with no additional content or external constraint.
full rationale
The paper is explicitly theoretical and defines the internal-time coordinates θ(t) and Tσ(t) via integrals over presupposed functions f(s) and σz(s), then sets APBTE(t) = Tσ(t)/Nref as the normalized age. This makes the central claim that APBTE measures the consumed fraction of the reference budget true by definitional construction rather than by independent derivation or external benchmark. The manuscript supplies no mapping from observables or closure relation that would make the coordinate falsifiable outside the definitions themselves. No load-bearing self-citation chain or fitted prediction is exhibited beyond the definitional step.
Axiom & Free-Parameter Ledger
free parameters (2)
- sref
- Nref
axioms (1)
- domain assumption Principle of Biological Time Equivalence (PBTE)
invented entities (3)
-
internal-time coordinate θ(t)
no independent evidence
-
entropy-normalized time Tσ(t)
no independent evidence
-
normalized PBTE age APBTE(t)
no independent evidence
Reference graph
Works this paper leans on
-
[1]
The hallmarks of aging,
C. López-Otín, M. A. Blasco, L. Partridge, M. Serrano, and G. Kroemer, “The hallmarks of aging,”Cell153, 1194–1217 (2013)
2013
-
[2]
Hallmarks of aging: An expanding universe,
C. López-Otín, M. A. Blasco, L. Partridge, M. Serrano, and G. Kroemer, “Hallmarks of aging: An expanding universe,”Cell186, 243–278 (2023)
2023
-
[3]
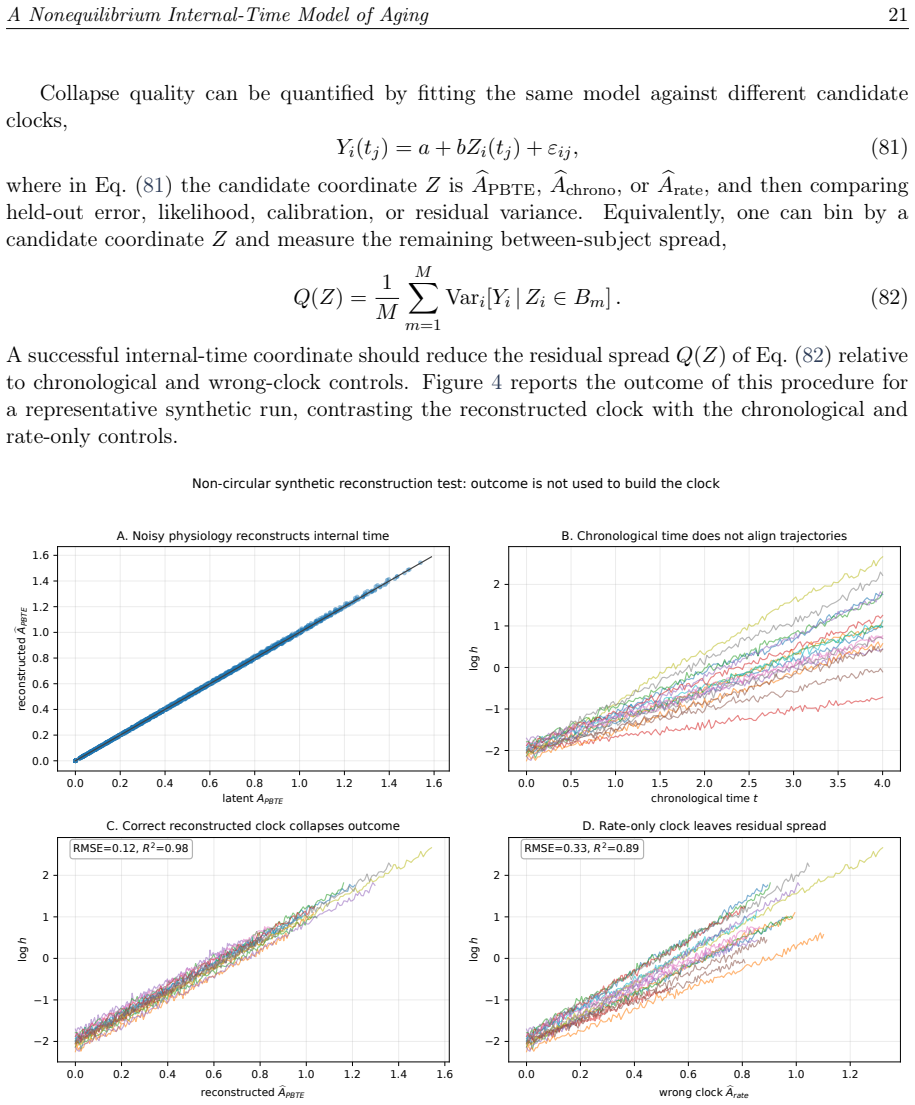
Biological proper time and entropy-cost invariance in cardiac and respiratory lifespan scaling
M. A. Taye, “Biological proper time and entropy-cost invariance in cardiac and respiratory lifespan scaling,” arXiv preprint arXiv:2606.15310 (2026)
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[4]
M. A. Taye,The Principle of Biological Time Equivalence: A Unified Theory of Life’s Temporal Invariants(book, 2026). A theoretical framework connecting biological time, metabolism, aging, physiological cycles, and entropy production
2026
-
[5]
M. A. Taye, “Biological Time Equivalence in Vertebrates: Thermodynamic Framework, Comparative Tests, and Clade-Specific Deviations,”Int. J. Sci. Res. Publ.16(5), 2250 (2026). arXiv:2603.26377
-
[6]
M. A. Taye, “The Lifetime Cardiac-Cycle Invariant in Endothermic Vertebrates: A 230- Species Comparative Dataset, Statistical Validation, and Explicit Falsifiability Criteria,” Int. J. Sci. Res. Publ.16(5), 2250 (2026). arXiv:2604.27856
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[7]
Thermodynamic Parametrisation of the Vertebrate Lifetime Cycle Invariant: BiologicalProperTime, AllometricMass-Cancellation, andClade-SpecificPredictions,
M. A. Taye, “Thermodynamic Parametrisation of the Vertebrate Lifetime Cycle Invariant: BiologicalProperTime, AllometricMass-Cancellation, andClade-SpecificPredictions,”Int. J. Sci. Res. Publ.16(4), 2250 (2026)
2026
-
[8]
M. A. Taye, “Neural Investment as an Entropy-Budget Strategy: A Thermodynamic Deriva- tion of Primate Longevity from the Principle of Biological Time Equivalence,” arXiv preprint arXiv:2604.27937 (2026)
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[9]
A general model for the origin of allometric scaling laws in biology,
G. B. West, J. H. Brown, and B. J. Enquist, “A general model for the origin of allometric scaling laws in biology,”Science276, 122–126 (1997). A Nonequilibrium Internal-Time Model of Aging26
1997
-
[10]
Schmidt-Nielsen,Scaling: Why Is Animal Size So Important?(Cambridge University Press, 1984)
K. Schmidt-Nielsen,Scaling: Why Is Animal Size So Important?(Cambridge University Press, 1984)
1984
-
[11]
Life and death: metabolic rate, membrane composition, and life span of animals,
A. J. Hulbert, R. Pamplona, R. Buffenstein, and W. A. Buttemer, “Life and death: metabolic rate, membrane composition, and life span of animals,”Physiol. Rev.87, 1175– 1213 (2007)
2007
-
[12]
C. E. Finch,Longevity, Senescence, and the Genome(University of Chicago Press, 1990)
1990
-
[13]
Localization at complex I and mechanism of the higher free radical production of brain nonsynaptic mitochondria in the short-lived rat,
G. Barja and A. Herrero, “Localization at complex I and mechanism of the higher free radical production of brain nonsynaptic mitochondria in the short-lived rat,”J. Bioenerg. Biomembr.30, 235–243 (1998)
1998
-
[14]
On the nature of the function expressive of the law of human mortality,
B. Gompertz, “On the nature of the function expressive of the law of human mortality,” Phil. Trans. R. Soc. Lond.115, 513–583 (1825)
-
[15]
Deciphering death: A commentary on Gompertz (1825),
T. B. L. Kirkwood, “Deciphering death: A commentary on Gompertz (1825),”Phil. Trans. R. Soc. B370, 20140379 (2015)
2015
-
[16]
Caloric restriction delays disease onset and mortality in rhesus monkeys,
R. J. Colman, R. M. Anderson, S. C. Johnson,et al., “Caloric restriction delays disease onset and mortality in rhesus monkeys,”Science325, 201–204 (2009)
2009
-
[17]
Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys,
R. J. Colman, T. M. Beasley, J. W. Kemnitz, S. C. Johnson, R. Weindruch, and R. M. An- derson, “Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys,” Nat. Commun.5, 3557 (2014)
2014
-
[18]
Caloric restriction improves health and survival of rhesus monkeys,
J. A. Mattison, R. J. Colman, T. M. Beasley,et al., “Caloric restriction improves health and survival of rhesus monkeys,”Nat. Commun.8, 14063 (2017)
2017
-
[19]
Mammalian hibernation: cellular and molecular responses to depressed metabolism and low temperature,
H. V. Carey, M. T. Andrews, and S. L. Martin, “Mammalian hibernation: cellular and molecular responses to depressed metabolism and low temperature,”Physiol. Rev.83, 1153– 1181 (2003)
2003
-
[20]
Schrödinger,What Is Life?(Cambridge University Press, 1944)
E. Schrödinger,What Is Life?(Cambridge University Press, 1944)
1944
-
[21]
Prigogine,Introduction to Thermodynamics of Irreversible Processes, 3rd ed
I. Prigogine,Introduction to Thermodynamics of Irreversible Processes, 3rd ed. (Wiley, 1967)
1967
-
[22]
Stochastic thermodynamics, fluctuation theorems and molecular machines,
U. Seifert, “Stochastic thermodynamics, fluctuation theorems and molecular machines,” Rep. Prog. Phys.75, 126001 (2012)
2012
-
[23]
M. A. Taye,Nonequilibrium Thermodynamics in Stochastic Processes: From Langevin Dy- namics to Brownian Motors(book, 2026). Develops the physical foundations of stochas- tic thermodynamics, Langevin dynamics, Brownian motion, entropy production, and fluctuation-driven transport
2026
-
[24]
Entropy Production from Macroscopic Balances: Bridging Continuum Laws and Stochastic Dynamics,
M. A. Taye, “Entropy Production from Macroscopic Balances: Bridging Continuum Laws and Stochastic Dynamics,”Mod. Math. Phys.2(1), 1 (2026)
2026
-
[25]
Hallmarks of cancer: the next generation,
D. Hanahan and R. A. Weinberg, “Hallmarks of cancer: the next generation,”Cell144, 646–674 (2011)
2011
-
[26]
Understanding the Warburg effect: the metabolic requirements of cell proliferation,
M. G. Vander Heiden, L. C. Cantley, and C. B. Thompson, “Understanding the Warburg effect: the metabolic requirements of cell proliferation,”Science324, 1029–1033 (2009)
2009
-
[27]
Digital health: tracking physiomes and activity using wearable biosensors,
X. Li, J. Dunn, D. Salins,et al., “Digital health: tracking physiomes and activity using wearable biosensors,”npj Digit. Med.1, 18 (2017)
2017
-
[28]
Universal DNA methylation age across mammalian tissues,
A. T. Lu, Z. Fei, A. Haghani,et al., “Universal DNA methylation age across mammalian tissues,”Nat. Aging3, 1148–1166 (2023). A Nonequilibrium Internal-Time Model of Aging27
2023
-
[29]
DNA methylation age of human tissues and cell types,
S. Horvath, “DNA methylation age of human tissues and cell types,”Genome Biol.14, R115 (2013)
2013
-
[30]
An epigenetic biomarker of aging for lifespan and healthspan,
M. E. Levine, A. T. Lu, A. Quach,et al., “An epigenetic biomarker of aging for lifespan and healthspan,”Aging10, 573–591 (2018)
2018
-
[31]
DNA methylation GrimAge strongly predicts lifespan and healthspan,
A. T. Lu, A. Quach, J. G. Wilson,et al., “DNA methylation GrimAge strongly predicts lifespan and healthspan,”Aging11, 303–327 (2019)
2019
-
[32]
A. T. Winfree,The Geometry of Biological Time(Springer, 1980)
1980
-
[33]
It’s all in the timing: the circadian clock in healthy aging,
S. Panda and J. B. Hogenesch, “It’s all in the timing: the circadian clock in healthy aging,” Nat. Rev. Immunol.21, 269–279 (2021)
2021
-
[34]
The free-energy principle: a unified brain theory?
K. Friston, “The free-energy principle: a unified brain theory?”Nat. Rev. Neurosci.11, 127–138 (2010)
2010
-
[35]
Scaling of brain metabolism with a fixed energy budget per neuron,
S. Herculano-Houzel, “Scaling of brain metabolism with a fixed energy budget per neuron,” PLoS ONE6, e17514 (2011)
2011
-
[36]
M. A. Taye,Brownian Motors and Brownian Heat Engines: From Classical Thermodynam- ics to Fluctuation-Driven Machines(book, 2026). Focuses on Brownian motors, Brownian heat engines, fluctuation-driven machines, rectification, transport, and thermodynamic ef- ficiency in small stochastic systems
2026
-
[37]
Exact Thermodynamic Analysis of a Hybrid Molecular Motor Switching Be- tween Active and Passive Modes,
M. A. Taye, “Exact Thermodynamic Analysis of a Hybrid Molecular Motor Switching Be- tween Active and Passive Modes,”Int. J. Sci. Res. Publ.16(5), 2250 (2026)
2026
-
[38]
A Universal Thermodynamic Inequality: Scaling Relations Between Current, Activity, and Entropy Production,
M. A. Taye, “A Universal Thermodynamic Inequality: Scaling Relations Between Current, Activity, and Entropy Production,”Int. J. Sci. Res. Publ.16(5), 2250 (2026)
2026
-
[39]
Noise-Activated Dopant Dynamics in Two-Dimensional Thermal Landscapes with Localized Cold Spots,
M. A. Taye, “Noise-Activated Dopant Dynamics in Two-Dimensional Thermal Landscapes with Localized Cold Spots,”Int. J. Sci. Res. Publ.16(5), 2250 (2026)
2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.