Activity-dependent neuromodulation and calcium homeostasis cooperate to produce robust and modulable neuronal function
Pith reviewed 2026-05-23 08:22 UTC · model grok-4.3
The pith
Calcium homeostasis and activity-dependent neuromodulation cooperate when their targets intersect in conductance space.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
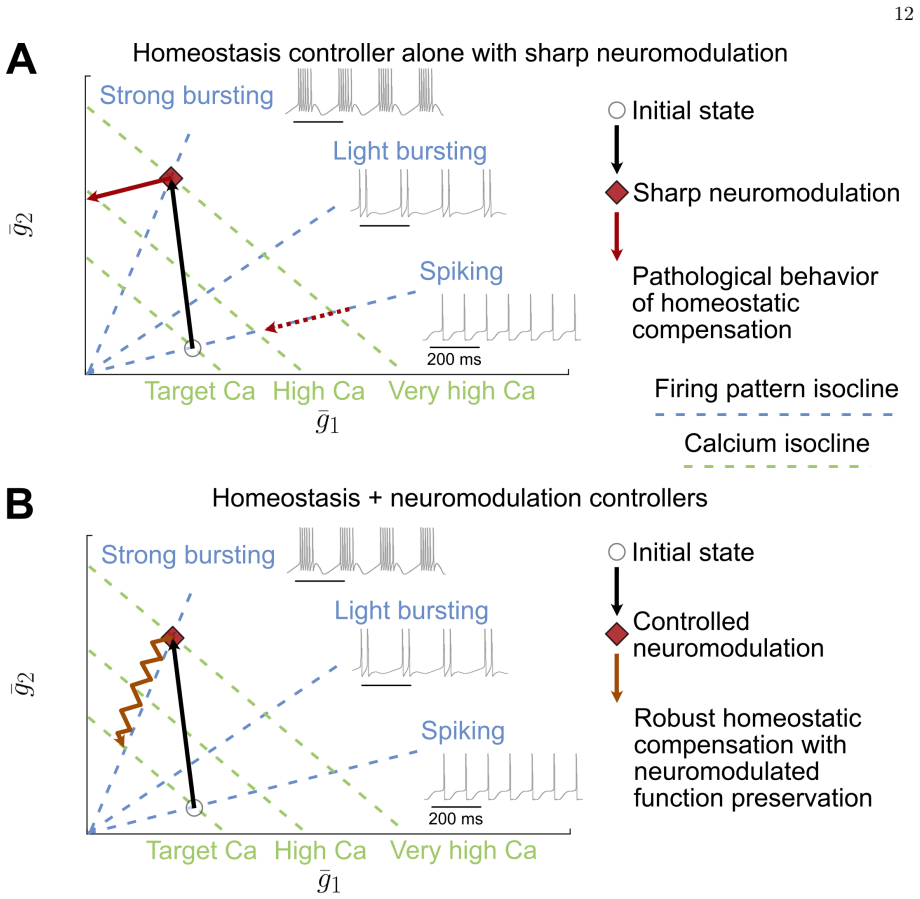
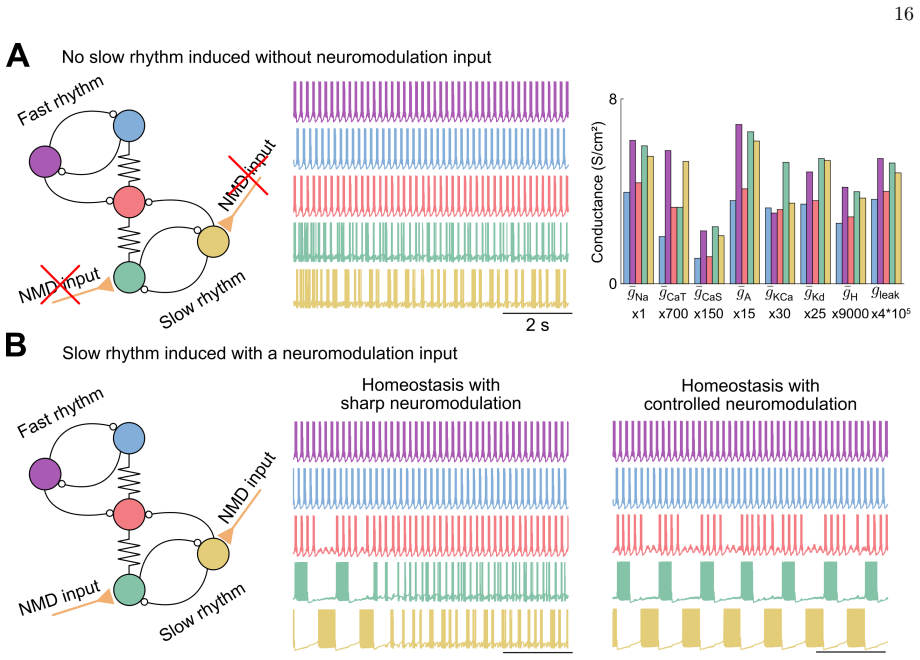
Using computational models of stomatogastric ganglion and dopaminergic neurons, the study demonstrates that a controlled neuromodulation mechanism preserves neuronal firing patterns while calcium homeostasis simultaneously maintains target intracellular calcium levels. Unlike sharp neuromodulation, the activity-dependent controller integrates feedback through mechanisms mimicking G-protein-coupled receptor cascades. The interaction between the controllers critically depends on the existence of an intersection in conductance space that balances target calcium levels against neuromodulated firing patterns, and maximizing neuronal degeneracy increases the likelihood of such intersections.
What carries the argument
intersection in conductance space between the calcium-homeostasis target and the neuromodulation-controlled firing pattern
If this is right
- Preserves firing patterns while calcium homeostasis maintains target levels
- Enables compensation for channel blockades when degeneracy is high
- Extends to reliable modulation of rhythmic activity in central pattern generators
- Supports robust and adaptive neural activity under physiological and pathological conditions
Where Pith is reading between the lines
- The same intersection requirement may apply to other pairs of homeostatic and modulatory controllers in different neuron classes.
- Disruption of degeneracy could be tested as a contributor to loss of robustness in models of channel-related pathologies.
- Network simulations could check whether the single-cell intersection principle scales to coordinated modulation across multiple cells.
- Experimental measurement of conductance trajectories during natural neuromodulation could confirm whether real neurons traverse or avoid the predicted intersection region.
Load-bearing premise
An intersection must exist in conductance space between the calcium-homeostasis target and the neuromodulation-controlled firing pattern.
What would settle it
A model run or experiment in which the two controllers are combined yet no conductance set simultaneously satisfies both the target calcium level and the desired firing pattern, resulting in either unstable calcium or loss of the firing pattern.
Figures




read the original abstract
Neurons rely on two interdependent mechanisms, homeostasis and neuromodulation, to maintain robust and adaptable functionality. Calcium homeostasis stabilizes neuronal activity by adjusting ionic conductances, whereas neuromodulation dynamically modifies ionic properties in response to external signals carried by neuromodulators. Combining these mechanisms in conductance-based models often produces unreliable outcomes, particularly when sharp neuromodulation interferes with calcium-homeostatic tuning. This study explores how a biologically inspired neuromodulation controller can harmonize with calcium homeostasis to ensure reliable neuronal function. Using computational models of stomatogastric ganglion and dopaminergic neurons, we demonstrate that controlled neuromodulation preserves neuronal firing patterns while calcium homeostasis simultaneously maintains target intracellular calcium levels. Unlike sharp neuromodulation, the neuromodulation controller integrates activity-dependent feedback through mechanisms mimicking G-protein-coupled receptor cascades. The interaction between these controllers critically depends on the existence of an intersection in conductance space, representing a balance between target calcium levels and neuromodulated firing patterns. Maximizing neuronal degeneracy enhances the likelihood of such intersections, enabling robust modulation and compensation for channel blockades. We further show that this controller pairing extends to network-level activity, reliably modulating the rhythmic activity of central pattern generators. This study highlights the complementary roles of calcium homeostasis and neuromodulation, proposing a unified control framework for maintaining robust and adaptive neural activity under physiological and pathological conditions.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that activity-dependent neuromodulation (implemented via GPCR-mimicking feedback) and calcium homeostasis can be combined in conductance-based models of stomatogastric ganglion (STG) and dopaminergic neurons to produce reliable firing patterns while maintaining target intracellular calcium; this harmonization requires an intersection in conductance space between the neuromodulation-controlled firing target and the calcium-homeostasis target, with neuronal degeneracy increasing the probability of such intersections and enabling compensation for channel blockades; the framework is further shown to modulate rhythmic activity in central pattern generator networks.
Significance. If the conductance-space intersection is explicitly located and shown to be robust across the reported models, the work would offer a concrete mechanistic account of how two canonical control systems can be made complementary rather than antagonistic, with direct implications for understanding degeneracy-enabled robustness and for modeling pathological disruptions of either controller.
major comments (2)
- [Abstract] Abstract: the central claim that 'the interaction between these controllers critically depends on the existence of an intersection in conductance space' is load-bearing for the harmonization result, yet the abstract (and by extension the reported simulations) provides no explicit demonstration—via nullcline analysis, fixed-point search, or systematic parameter sweeps over the free conductance parameters—that such an intersection is attained rather than assumed for the STG and dopaminergic models.
- [Abstract] Abstract: the assertion that 'maximizing neuronal degeneracy enhances the likelihood of such intersections' is presented as enabling robust modulation and compensation, but no quantitative comparison (e.g., intersection frequency in high- vs. low-degeneracy regimes or sensitivity analysis on the listed conductance parameters) is described to support this dependence.
minor comments (1)
- [Abstract] Abstract: 'sharp neuromodulation' is contrasted with the proposed controller but is never defined; a brief operational definition or reference to the implementation difference would improve clarity.
Simulated Author's Rebuttal
We thank the referee for their constructive comments, which highlight opportunities to strengthen the explicit demonstration of key claims in our work. We address each major comment below and will incorporate revisions to provide the requested analyses.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that 'the interaction between these controllers critically depends on the existence of an intersection in conductance space' is load-bearing for the harmonization result, yet the abstract (and by extension the reported simulations) provides no explicit demonstration—via nullcline analysis, fixed-point search, or systematic parameter sweeps over the free conductance parameters—that such an intersection is attained rather than assumed for the STG and dopaminergic models.
Authors: We agree that an explicit demonstration of the conductance-space intersection would strengthen the manuscript. While our simulations show convergence to stable states satisfying both target calcium and firing-pattern criteria, we did not include nullcline analysis, fixed-point searches, or systematic sweeps over free conductances. We will revise the manuscript to add these, including a new figure with conductance-space target lines and their intersections for both neuron models. revision: yes
-
Referee: [Abstract] Abstract: the assertion that 'maximizing neuronal degeneracy enhances the likelihood of such intersections' is presented as enabling robust modulation and compensation, but no quantitative comparison (e.g., intersection frequency in high- vs. low-degeneracy regimes or sensitivity analysis on the listed conductance parameters) is described to support this dependence.
Authors: We acknowledge that the claim about degeneracy increasing intersection likelihood is currently supported by qualitative reasoning on parameter-space dimensionality rather than quantitative evidence. We will add comparative analyses in the revision, such as Monte Carlo sampling across high- and low-degeneracy conductance regimes to report intersection frequencies and sensitivity to parameter variation. revision: yes
Circularity Check
No significant circularity; central claim demonstrated via model simulations rather than reduced to inputs by construction.
full rationale
The paper presents computational models of stomatogastric ganglion and dopaminergic neurons to show that activity-dependent neuromodulation (via GPCR-mimicking feedback) harmonizes with calcium homeostasis when an intersection exists in conductance space. This intersection is treated as an emergent property explored through the models, not defined into existence or fitted to guarantee the outcome. No load-bearing self-citations, uniqueness theorems from prior author work, or ansatzes smuggled via citation are evident in the abstract or described approach. The result relies on explicit simulation of controller interactions and degeneracy effects, which are independently falsifiable in the chosen neuron models. This is the most common honest finding for model-based studies that do not rename known results or call fits predictions.
Axiom & Free-Parameter Ledger
free parameters (1)
- conductance parameters in stomatogastric and dopaminergic models
axioms (1)
- domain assumption Standard Hodgkin-Huxley-style conductance equations accurately capture the relevant dynamics of the modeled neurons.
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The interaction between these controllers critically depends on the existence of an intersection in conductance space, representing a balance between target calcium levels and neuromodulated firing patterns
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Maximizing neuronal degeneracy enhances the likelihood of such intersections
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Forward citations
Cited by 1 Pith paper
-
Neuromodulation supports robust rhythmic pattern transitions in degenerate central pattern generators with fixed connectivity
An adaptive neuromodulation controller using equivariant bifurcation theory enables robust gait transitions in degenerate central pattern generators with fixed connectivity.
Reference graph
Works this paper leans on
-
[1]
Anger, T., Madge, D. J., Mulla, M., and Riddall, D. (2001). Medicinal chemistry of neuronal voltage-gated sodium channel blockers. Journal of Medicinal Chemistry , 44(2):115–137
work page 2001
-
[2]
Bargmann, C. I. and Marder, E. (2013). From the connectome to brain function. Nature methods, 10(6):483–490
work page 2013
-
[3]
Bezanson, J., Edelman, A., Karpinski, S., and Shah, V. B. (2017). Julia: A fresh approach to numerical computing. SIAM review, 59(1):65–98
work page 2017
-
[4]
Cunha, R. (2001). Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: Different roles, different sources and different receptors. Neurochemistry International, 38(2):107–125
work page 2001
-
[5]
Desai, N. S. (2003). Homeostatic plasticity in the cns: synaptic and intrinsic forms. Journal of Physiology- Paris, 97(4-6):391–402
work page 2003
-
[6]
DeWitt, C. R. and Waksman, J. C. (2004). Pharmacology, pathophysiology and management of calcium channel blocker and β-blocker toxicity. Toxicological Reviews, 23:223–238
work page 2004
-
[7]
Drion, G., Franci, A., and Sepulchre, R. (2019). Cellular switches orchestrate rhythmic circuits.Biological cybernetics, 113:71–82
work page 2019
-
[8]
Drion, G., Massotte, L., Sepulchre, R., and Seutin, V. (2011). How modeling can reconcile apparently discrepant experimental results: the case of pacemaking in dopaminergic neurons. PLoS Computational Biology, 7(5):e1002050
work page 2011
-
[9]
Fyon, A., Franci, A., Sacr´ e, P., and Drion, G. (2024). Dimensionality reduction of neuronal degeneracy reveals two interfering physiological mechanisms. PNAS nexus, 3(10):pgae415
work page 2024
-
[10]
Fyon, A., Sacr´ e, P., Franci, A., and Drion, G. (2023). Reliable neuromodulation from adaptive control of ion channel expression. IFAC-PapersOnLine, 56(2):458–463
work page 2023
-
[11]
Goaillard, J.-M. and Marder, E. (2021). Ion channel degeneracy, variability, and covariation in neuron and circuit resilience. Annual review of neuroscience, 44:335–357
work page 2021
-
[12]
Grant, A., Starmer, C., and Strauss, H. (1984). Antiarrhythmic drug action. blockade of the inward sodium current. Circulation Research, 55(4):427–439
work page 1984
-
[13]
Guertin, P. A. (2009). The mammalian central pattern generator for locomotion. Brain research reviews, 62(1):45–56
work page 2009
-
[14]
J., O’Leary, T., and Marder, E
Gutierrez, G. J., O’Leary, T., and Marder, E. (2013). Multiple mechanisms switch an electrically coupled, synaptically inhibited neuron between competing rhythmic oscillators. Neuron, 77(5):845–858
work page 2013
-
[15]
Hodgkin, A. L. and Huxley, A. F. (1952). A quantitative description of membrane current and its application to conduction and excitation in nerve. The Journal of physiology , 117(4):500
work page 1952
-
[16]
Hooper, S. L. and Marder, E. (1984). Modulation of a central pattern generator by two neuropeptides, proctolin and fmrfamide. Brain research, 305(1):186–191
work page 1984
-
[17]
Imbrici, P., Nicolotti, O., Leonetti, F., Conte, D., and Liantonio, A. (2018). Ion channels in drug discovery and safety pharmacology. Computational Toxicology: Methods and Protocols , pages 313–326
work page 2018
-
[18]
Izhikevich, E. M. (2007). Dynamical systems in neuroscience . MIT press
work page 2007
-
[19]
J., Onizuka, M., Kawato, M., and Aihara, K
Katori, Y., Lang, E. J., Onizuka, M., Kawato, M., and Aihara, K. (2010). Quantitative modeling of spatio- temporal dynamics of inferior olive neurons with a simple conductance-based model. International Journal of Bifurcation and Chaos , 20(03):583–603
work page 2010
-
[20]
Kramer, R. H. and Levitan, I. B. (1990). Activity-dependent neuromodulation in aplysia neuron r15: intracellular calcium antagonizes neurotransmitter responses mediated by camp. Journal of neurophysiology, 63(5):1075–1088. 21
work page 1990
-
[21]
Liu, Z., Golowasch, J., Marder, E., and Abbott, L. (1998). A model neuron with activity-dependent conductances regulated by multiple calcium sensors. Journal of Neuroscience, 18(7):2309–2320
work page 1998
-
[22]
Marcus, D. J. and Bruchas, M. R. (2023). Optical approaches for investigating neuromodulation and g protein–coupled receptor signaling. Pharmacological Reviews, 75(6):1119–1139
work page 2023
-
[23]
Marder, E. (2011). Variability, compensation, and modulation in neurons and circuits. Proceedings of the National Academy of Sciences , 108(supplement 3):15542–15548
work page 2011
-
[24]
Marder, E. (2012). Neuromodulation of neuronal circuits: back to the future. Neuron, 76(1):1–11
work page 2012
-
[25]
Marder, E. and Bucher, D. (2001). Central pattern generators and the control of rhythmic movements. Current biology, 11(23):R986–R996
work page 2001
-
[26]
Marder, E. and Bucher, D. (2005). Robustness in neuronal systems: The balance between homeostasis, plasticity, and modulation. In Robust design: A repertoire of biological, ecological and engineering case studies. Oxford University Press Oxford
work page 2005
-
[27]
Marder, E. and Calabrese, R. L. (1996). Principles of rhythmic motor pattern generation. Physiological reviews, 76(3):687–717
work page 1996
-
[28]
Marder, E. and Goaillard, J.-M. (2006). Variability, compensation and homeostasis in neuron and network function. Nature Reviews Neuroscience, 7(7):563–574
work page 2006
-
[29]
Marder, E., O’Leary, T., and Shruti, S. (2014). Neuromodulation of circuits with variable parameters: single neurons and small circuits reveal principles of state-dependent and robust neuromodulation. Annual review of neuroscience, 37:329–346
work page 2014
-
[30]
Marder, E. and Prinz, A. A. (2002). Modeling stability in neuron and network function: the role of activity in homeostasis. Bioessays, 24(12):1145–1154
work page 2002
-
[31]
McCormick, D. A., Nestvogel, D. B., and He, B. J. (2020). Neuromodulation of brain state and behavior. Annual review of neuroscience, 43:391–415
work page 2020
-
[32]
Megwa, O. F., Pascual, L. M., G¨ unay, C., Pulver, S. R., and Prinz, A. A. (2023). Temporal dynamics of na/k pump mediated memory traces: insights from conductance-based models of drosophila neurons. Frontiers in Neuroscience, 17:1154549
work page 2023
-
[33]
Nadim, F. and Bucher, D. (2014). Neuromodulation of neurons and synapses. Current opinion in neurobiology, 29:48–56. O’Leary, T., Williams, A. H., Caplan, J. S., and Marder, E. (2013). Correlations in ion channel expression emerge from homeostatic tuning rules. Proceedings of the National Academy of Sciences , 110(28):E2645–E2654. O’Leary, T., van Rossum,...
work page 2014
-
[34]
Plant, R. E. (1981). Bifurcation and resonance in a model for bursting nerve cells.Journal of mathematical biology, 11:15–32
work page 1981
-
[35]
Pratt, K. G. and Aizenman, C. D. (2007). Homeostatic regulation of intrinsic excitability and synaptic transmission in a developing visual circuit. Journal of Neuroscience, 27(31):8268–8277
work page 2007
-
[36]
Prinz, A. A. (2017). Degeneracy rules! The Journal of Physiology , 595(8):2409
work page 2017
-
[37]
Qian, K., Yu, N., Tucker, K. R., Levitan, E. S., and Canavier, C. C. (2014). Mathematical analysis of depolarization block mediated by slow inactivation of fast sodium channels in midbrain dopamine neurons. Journal of neurophysiology, 112(11):2779–2790
work page 2014
-
[38]
Rathour, R. K. and Narayanan, R. (2019). Degeneracy in hippocampal physiology and plasticity. Hippocampus, 29(10):980–1022. 22
work page 2019
-
[39]
Raymond, J. L., Baxter, D. A., Buonomano, D. V., and Byrne, J. H. (1992). A learning rule based on empirically-derived activity-dependent neuromodulation supports operant conditioning in a small network. Neural Networks, 5(5):789–803
work page 1992
-
[40]
Scheler, G. (2004). Regulation of neuromodulator receptor efficacy—implications for whole-neuron and synaptic plasticity. Progress in Neurobiology, 72(6):399–415
work page 2004
-
[41]
Selverston, A., Sz¨ ucs, A., Huerta, R., Pinto, R. D., and Reyes, M. B. (2009). Neural mechanisms underlying the generation of the lobster gastric mill motor pattern. Frontiers in neural circuits, 3:608
work page 2009
-
[42]
Starmer, C. F., Lastra, A. A., Nesterenko, V. V., and Grant, A. O. (1991). Proarrhythmic response to sodium channel blockade. theoretical model and numerical experiments. Circulation, 84(3):1364–1377
work page 1991
-
[43]
Wechselberger, M., Wright, C. L., Bishop, G. A., and Boulant, J. A. (2006). Ionic channels and conductance-based models for hypothalamic neuronal thermosensitivity. American Journal of
work page 2006
-
[44]
Werner, G. and Mitterauer, B. J. (2014). The dynamics of neuromodulation. Criticality in Neural Systems, pages 525–538
work page 2014
-
[45]
Whitacre, J. and Bender, A. (2010). Degeneracy: A design principle for achieving robustness and evolvability. Journal of Theoretical Biology, 263(1):143–153
work page 2010
-
[46]
Whitacre, J. M. (2010). Degeneracy: A link between evolvability, robustness and complexity in biological systems. Theoretical Biology and Medical Modelling, 7:1–17. X Cubeddu, L. (2016). Drug-induced inhibition and trafficking disruption of ion channels: Pathogenesis of qt abnormalities and drug-induced fatal arrhythmias. Current Cardiology Reviews, 12(2):141–154
work page 2010
-
[47]
Yuste, R., MacLean, J. N., Smith, J., and Lansner, A. (2005). The cortex as a central pattern generator. Nature Reviews Neuroscience, 6(6):477–483
work page 2005
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.