A Mechanistic Model for Collective Motion from Sensorimotor Regularities
Pith reviewed 2026-05-20 17:29 UTC · model grok-4.3
The pith
Collective motion in animals emerges when agents keep desired distances using only bearing and apparent-size cues from neighbors.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Each agent perceives neighbors through bearing and apparent-size cues within a limited field of view, maintains uncertain internal state estimates, and selects actions through gradient descent on a desired social distance without any prescribed interaction forces. This simple model produces diverse collective behaviors including polarized motion, milling, ring formations, and subgroup fragmentation. A global sensitivity analysis shows that behavioral transitions are governed by sensorimotor parameters corresponding to measurable biological quantities: field of view geometry, sensory noise, turning agility, and memory.
What carries the argument
Gradient descent on a desired social distance computed from uncertain estimates of neighbor bearing and apparent size.
If this is right
- Diverse group patterns such as polarization, milling, rings, and fragmentation arise without any explicit alignment or attraction forces.
- Shifts between behaviors are produced by changes in field-of-view angle, sensory noise, turning agility, and memory duration.
- Differences in collective motion across species follow from differences in sensorimotor parameters tied to body size and habitat.
Where Pith is reading between the lines
- Empirical work could measure how altering an animal's effective field of view changes group cohesion and test the predicted transition thresholds.
- The same perception-action loop could be ported to robot swarms to achieve similar behaviors with minimal communication.
- Adding other sensory channels such as relative velocity or odor would likely expand the range of stable formations the model can produce.
Load-bearing premise
Agents maintain uncertain internal state estimates of neighbor positions and select actions exclusively through gradient descent on a desired social distance derived from bearing and apparent-size cues.
What would settle it
If locusts or similar animals are shown to match neighbor velocities directly rather than modulating their own speed or turn rate to hold an apparent-size distance, the model's action-selection rule would be falsified.
Figures




read the original abstract
Collective behavior in animals has long been modeled through self-propelled particle models, which reproduce striking group-level phenomena through abstract interaction forces. Yet these models are fundamentally descriptive: they leave open the question of how collective behavior is actually produced. Recent empirical work makes this gap concrete: locusts do not align with neighbors, sensory and cognitive mechanisms mediate interaction instead. A mechanistic model must therefore operate at the sensorimotor level, grounded in what individual organisms can actually perceive, estimate, and physically execute. We present such a model based on a modeling framework from robotics, extended here to collective motion. Each agent perceives neighbors through bearing and apparent-size cues within a limited field of view, maintains uncertain internal state estimates, and selects actions through gradient descent on a desired social distance -- without any prescribed interaction forces. This simple model produces diverse collective behaviors including polarized motion, milling, ring formations, and subgroup fragmentation. A global sensitivity analysis shows that behavioral transitions are governed by sensorimotor parameters corresponding to measurable biological quantities: field of view geometry, sensory noise, turning agility, and memory. Collective behavior can therefore be understood as the emergent outcome of interacting sensorimotor regularities, and differences across species as the emergent outcome of differences in embodiment and environment.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper presents a mechanistic model for collective motion in which agents perceive neighbors only through bearing and apparent-size cues within a limited field of view, maintain uncertain internal state estimates of neighbor positions, and select actions exclusively by gradient descent on a target social distance. No explicit alignment, attraction, or repulsion forces are prescribed. Simulations of this sensorimotor rule set are reported to produce polarized motion, milling, ring formations, and subgroup fragmentation. A global sensitivity analysis is used to show that transitions among these behaviors are controlled by parameters (field-of-view angle, sensory noise, turning agility, memory) that map to measurable biological quantities.
Significance. If the central claim holds, the work supplies a concrete alternative to descriptive self-propelled-particle models by grounding collective phenomena in explicit perception-action loops. The global sensitivity analysis that ties behavioral transitions to embodiment and environment parameters is a clear strength, as is the absence of post-hoc fitting to observed group statistics. The approach therefore offers a falsifiable, robotics-derived route to explaining both within-species collective patterns and cross-species differences.
major comments (1)
- [action selection mechanism] Action-selection section: the manuscript must specify the exact filter or Bayesian update rule used to maintain the uncertain internal state estimates of neighbor bearing and apparent size, and must state whether the gradient is evaluated on the instantaneous cue or on the filtered estimate. Without this detail it remains possible that the combination of state estimation and gradient descent on social distance effectively reproduces the attraction/repulsion or alignment dynamics of classical SPP models, undermining the claim that the behaviors arise solely from sensorimotor regularities without prescribed interaction forces.
minor comments (2)
- Figure captions should explicitly label which panels correspond to which collective regime (polarized, milling, ring, fragmentation) and should indicate the parameter values used for each simulation.
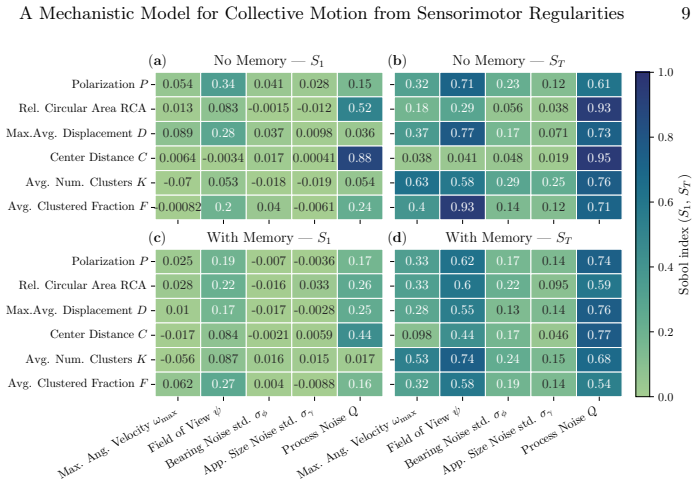
- The global sensitivity analysis would benefit from a brief statement of the sampling method (e.g., Sobol, Latin hypercube) and whether confidence intervals or error bars are reported for the sensitivity indices.
Simulated Author's Rebuttal
We thank the referee for the constructive review and positive assessment of the significance of our work. We address the single major comment below and will incorporate the requested clarifications in the revised manuscript.
read point-by-point responses
-
Referee: Action-selection section: the manuscript must specify the exact filter or Bayesian update rule used to maintain the uncertain internal state estimates of neighbor bearing and apparent size, and must state whether the gradient is evaluated on the instantaneous cue or on the filtered estimate. Without this detail it remains possible that the combination of state estimation and gradient descent on social distance effectively reproduces the attraction/repulsion or alignment dynamics of classical SPP models, undermining the claim that the behaviors arise solely from sensorimotor regularities without prescribed interaction forces.
Authors: We agree that explicit specification of the state-estimation procedure is required for reproducibility and to substantiate the mechanistic claim. In the revised manuscript we will add a dedicated subsection in the Methods that states: (i) the internal estimates are maintained via a discrete-time Kalman filter whose process and measurement noise covariances are set by the sensory-noise parameter; (ii) the filter updates both bearing and apparent-size estimates from the instantaneous visual cues; and (iii) the gradient of social distance is evaluated exclusively on the filtered state estimates, not on the raw cues. The resulting control law therefore contains no explicit alignment, attraction or repulsion terms; any apparent similarity to classical SPP forces is an emergent consequence of the closed sensorimotor loop rather than an imposed interaction rule. We will also include the filter equations and a brief proof that the closed-loop dynamics differ from the standard Vicsek or Couzin formulations. revision: yes
Circularity Check
No circularity: collective patterns emerge from explicit sensorimotor rules in simulation
full rationale
The paper constructs an explicit mechanistic model in which agents perceive neighbors via bearing and apparent-size cues, maintain uncertain internal estimates, and select actions by gradient descent on a target social distance, without any prescribed forces or alignment rules. Collective behaviors (polarized motion, milling, etc.) are then generated as simulation outputs from these rules. This is a constructive forward simulation rather than a derivation in which the target phenomena are presupposed or fitted by construction. No load-bearing step reduces to a self-citation, a fitted parameter renamed as prediction, or an ansatz smuggled via prior work; the sensitivity analysis further treats the sensorimotor parameters as independent inputs whose effects on outputs are examined. The central claim therefore remains non-circular.
Axiom & Free-Parameter Ledger
free parameters (3)
- desired social distance
- field of view angle
- sensory noise level
axioms (2)
- domain assumption Agents can form uncertain internal estimates of neighbor positions from bearing and apparent-size cues alone.
- domain assumption Action selection occurs by gradient descent on the error between current and desired social distance.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel echoes?
echoesECHOES: this paper passage has the same mathematical shape or conceptual pattern as the Recognition theorem, but is not a direct formal dependency.
each agent ... selects actions through gradient descent on a desired social distance—without any prescribed interaction forces. This simple model produces diverse collective behaviors
-
IndisputableMonolith/Foundation/LogicAsFunctionalEquation.leanSatisfiesLawsOfLogic echoes?
echoesECHOES: this paper passage has the same mathematical shape or conceptual pattern as the Recognition theorem, but is not a direct formal dependency.
AICON generates behavior through the dynamic composition of sensorimotor regularities, without directly encoding behavior itself
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
PLoS Computational Biology10(7), e1003697 (2014)
Attanasi, A., Cavagna, A., Del Castello, L., Giardina, I., Melillo, S., Parisi, L., Pohl, O., Rossaro, B., Shen, E., Silvestri, E., et al.: Collective behaviour without collective order in wild swarms of midges. PLoS Computational Biology10(7), e1003697 (2014)
work page 2014
-
[2]
Proceedings of the National Academy of Sciences105(4), 1232– 1237 (2008)
Ballerini, M., Cabibbo, N., Candelier, R., Cavagna, A., Cisbani, E., Giardina, I., Lecomte, V., Orlandi, A., Parisi, G., Procaccini, A., et al.: Interaction ruling animal collective behavior depends on topological rather than metric distance: Evidence from a field study. Proceedings of the National Academy of Sciences105(4), 1232– 1237 (2008)
work page 2008
-
[3]
Science Advances6(6), eaay0792 (2020)
Bastien, R., Romanczuk, P.: A model of collective behavior based purely on vision. Science Advances6(6), eaay0792 (2020)
work page 2020
-
[4]
Buhl, C., Sumpter, D.J., Couzin, I.D., Hale, J.J., Despland, E., Miller, E.R., Simp- son,S.J.:Fromdisordertoorderinmarchinglocusts.Science312(5778),1402–1406 (2006)
work page 2006
-
[5]
Proceedings of the National Academy of Sciences107(26), 11865–11870 (2010)
Cavagna, A., Cimarelli, A., Giardina, I., Parisi, G., Santagati, R., Stefanini, F., Viale, M.: Scale-free correlations in starling flocks. Proceedings of the National Academy of Sciences107(26), 11865–11870 (2010)
work page 2010
-
[6]
Nature433(7025), 513–516 (2005)
Couzin, I.D., Krause, J., Franks, N.R., Levin, S.A.: Effective leadership and decision-making in animal groups on the move. Nature433(7025), 513–516 (2005)
work page 2005
-
[7]
Ginelli,F.:Thephysicsofthevicsekmodel.TheEuropeanPhysicalJournalSpecial Topics225(11), 2099–2117 (2016) 12 V. Mengers et al
work page 2099
-
[8]
Proceedings of the National Academy of Sciences121(17), e2320239121 (2024)
Heins, C., Millidge, B., Da Costa, L., Mann, R.P., Friston, K.J., Couzin, I.D.: Col- lective behavior from surprise minimization. Proceedings of the National Academy of Sciences121(17), e2320239121 (2024)
work page 2024
-
[9]
Proceedings of the National Academy of Sciences108(46), 18720–18725 (2011)
Katz,Y.,Tunstrøm,K.,Ioannou,C.C.,Huepe,C.,Couzin,I.D.:Inferringthestruc- ture and dynamics of interactions in schooling fish. Proceedings of the National Academy of Sciences108(46), 18720–18725 (2011)
work page 2011
-
[10]
Reviews of Modern Physics 85(3), 1143–1189 (2013)
Marchetti, M.C., Joanny, J.F., Ramaswamy, S., Liverpool, T.B., Prost, J., Rao, M., Simha, R.A.: Hydrodynamics of soft active matter. Reviews of Modern Physics 85(3), 1143–1189 (2013)
work page 2013
-
[11]
The International Journal of Robotics Research 41(8), 741–777 (2022)
Martín-Martín, R., Brock, O.: Coupled recursive estimation for online interactive perception of articulated objects. The International Journal of Robotics Research 41(8), 741–777 (2022)
work page 2022
-
[12]
In: IEEE International Conference on Robotics and Automation
Mengers, V., Brock, O.: No plan but everything under control: Robustly solving se- quential tasks with dynamically composed gradient descent. In: IEEE International Conference on Robotics and Automation. pp. 90–96 (2025)
work page 2025
-
[13]
Scientific Reports14, 27314 (2024)
Mengers, V., Raoufi, M., Brock, O., Hamann, H., Romanczuk, P.: Leveraging un- certainty in collective opinion dynamics with heterogeneity. Scientific Reports14, 27314 (2024)
work page 2024
-
[14]
Journal of Vision25(2), 6 (2025)
Mengers, V., Roth, N., Brock, O., Obermayer, K., Rolfs, M.: A robotics-inspired scanpath model reveals the importance of uncertainty and semantic object cues for gaze guidance in dynamic scenes. Journal of Vision25(2), 6 (2025)
work page 2025
-
[15]
PLoS Computational Biology20(5), e1012087 (2024)
Mezey, D., Deffner, D., Kurvers, R.H., Romanczuk, P.: Visual social information use in collective foraging. PLoS Computational Biology20(5), e1012087 (2024)
work page 2024
-
[16]
Proceedings of the National Academy of Sciences 112(15), 4690–4695 (2015)
Rosenthal, S.B., Twomey, C.R., Hartnett, A.T., Wu, H.S., Couzin, I.D.: Revealing the hidden networks of interaction in mobile animal groups allows prediction of complex behavioral contagion. Proceedings of the National Academy of Sciences 112(15), 4690–4695 (2015)
work page 2015
-
[17]
Nature Communications16(1), 9051 (2025)
Salahshour, M., Couzin, I.D.: Allocentric flocking. Nature Communications16(1), 9051 (2025)
work page 2025
-
[18]
Science387(6737), 995– 1000 (2025)
Sayin, S., Couzin-Fuchs, E., Petelski, I., Günzel, Y., Salahshour, M., Lee, C.Y., Graving, J.M., Li, L., Deussen, O., Sword, G.A., et al.: The behavioral mecha- nisms governing collective motion in swarming locusts. Science387(6737), 995– 1000 (2025)
work page 2025
-
[19]
ACM Transactions on Database Systems42(3), 19 (2017)
Schubert, E., Sander, J., Ester, M., Kriegel, H.P., Xu, X.: DBSCAN revisited, revisited: why and how you should (still) use DBSCAN. ACM Transactions on Database Systems42(3), 19 (2017)
work page 2017
-
[20]
Nature521(7551), 186–191 (2015)
Seelig, J.D., Jayaraman, V.: Neural dynamics for landmark orientation and angular path integration. Nature521(7551), 186–191 (2015)
work page 2015
-
[21]
Mathematics and Computers in Simulation55(1-3), 271– 280 (2001)
Sobol, I.M.: Global sensitivity indices for nonlinear mathematical models and their Monte Carlo estimates. Mathematics and Computers in Simulation55(1-3), 271– 280 (2001)
work page 2001
-
[22]
Current Biology 23(17), R709–R711 (2013)
Strandburg-Peshkin, A., Twomey, C.R., Bode, N.W., Kao, A.B., Katz, Y., Ioan- nou, C.C., Rosenthal, S.B., Torney, C.J., Wu, H.S., Levin, S.A., et al.: Visual sen- sory networks and effective information transfer in animal groups. Current Biology 23(17), R709–R711 (2013)
work page 2013
-
[23]
Princeton University Press (2010)
Sumpter, D.J.: Collective Animal Behavior. Princeton University Press (2010)
work page 2010
-
[24]
Physical Review Letters75(6), 1226 (1995)
Vicsek, T., Czirók, A., Ben-Jacob, E., Cohen, I., Shochet, O.: Novel type of phase transition in a system of self-driven particles. Physical Review Letters75(6), 1226 (1995)
work page 1995
-
[25]
Vicsek,T.,Zafeiris,A.:Collectivemotion.PhysicsReports517(3-4),71–140(2012)
work page 2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.