Recognition: unknown
Scale-dependent Temporal Signatures of Arboviral Transmission in Urban Environments
Pith reviewed 2026-05-10 16:10 UTC · model grok-4.3
The pith
Differences between dengue, Zika and chikungunya transmission emerge only at specific temporal scales rather than from spatial proximity.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
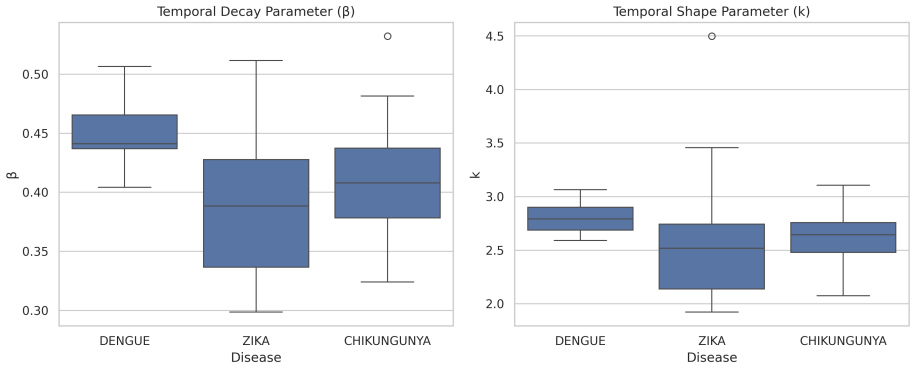
Interactions are expressed as a function of spatial distance and temporally delayed influence whose parameters are obtained by maximum likelihood. When fitted to the Recife data the spatial parameter collapses, indicating that proximity supplies no reliable separation among dengue, Zika and chikungunya. Temporal dynamics display scale dependence: statistical differentiation appears only beyond a minimum temporal window. Models without biological constraints mainly recover short-term co-occurrence and produce non-robust distinctions, whereas constrained models recover a shared underlying transmission structure. The resulting networks display localized, structured interaction patterns.
What carries the argument
Pairwise interaction kernel that encodes spatial distance and temporally delayed influence, with parameters estimated by maximum likelihood under biological constraints.
If this is right
- Unconstrained models primarily capture short-term co-occurrence and therefore generate apparent but non-robust differences between diseases.
- Biologically constrained models recover a common underlying transmission structure across the three arboviruses.
- Reconstructed transmission networks exhibit localized and structured interaction patterns consistent with plausible epidemic propagation.
- Epidemic differentiation is an emergent phenomenon that depends on the temporal scale at which the data are examined.
Where Pith is reading between the lines
- Public-health responses in dense cities may gain more from timing interventions to longer transmission windows than from purely spatial targeting.
- The same kernel framework could be applied to other urban datasets to test whether the critical temporal window length is conserved across cities.
- The collapse of spatial information raises the question of whether finer-scale mobility or vector-density layers would restore spatial discriminatory power.
Load-bearing premise
The pairwise interaction kernel with maximum-likelihood parameter estimation, once biologically constrained, correctly isolates the underlying transmission structure without missing unmodeled factors such as vector density variation or reporting biases.
What would settle it
Re-fitting the same model to the Recife data while removing the biological constraints and observing that temporal differentiation remains stable at all window lengths would falsify the claim that differentiation is scale-dependent.
Figures

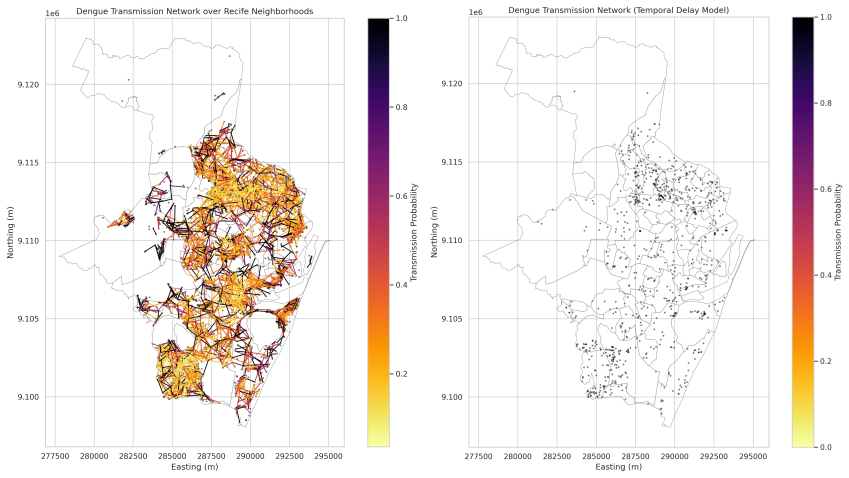


read the original abstract
Understanding epidemic dynamics in urban environments requires models that capture interactions across space and time while incorporating biological constraints. In this work, we propose a probabilistic spatiotemporal framework based on pairwise interaction kernels to analyze arboviral transmission using large-scale georeferenced data from Recife, Brazil. The model describes interactions as a function of spatial distance and temporally delayed influence, with parameters estimated via maximum likelihood. Our results reveal a marked asymmetry between spatial and temporal components. The spatial parameter systematically collapses, indicating that spatial proximity does not provide discriminatory information between diseases at the urban scale. In contrast, temporal dynamics exhibit scale-dependent behavior: statistical differentiation between dengue, Zika, and chikungunya emerges only beyond a critical temporal window. We show that unconstrained models primarily capture short-term co-occurrence, leading to apparent but non-robust differences, while biologically constrained models reveal a common underlying transmission structure. Additionally, reconstructed transmission networks exhibit localized and structured interaction patterns consistent with plausible epidemic propagation. These findings demonstrate that epidemic differentiation is not intrinsic, but an emergent phenomenon dependent on temporal scale, highlighting the importance of biologically grounded and scale-aware modeling in spatiotemporal epidemic analysis.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims to develop a probabilistic spatiotemporal framework using pairwise interaction kernels to model arboviral transmission in urban environments, applied to georeferenced case data from Recife, Brazil for dengue, Zika, and chikungunya. Key findings include the collapse of spatial parameters indicating lack of discriminatory information at urban scales, and scale-dependent temporal signatures where statistical differentiation between the diseases emerges only beyond a critical temporal window when biological constraints are applied. Unconstrained models are said to capture short-term co-occurrence leading to apparent differences, while constrained models reveal a common underlying structure, with reconstructed transmission networks showing localized patterns consistent with epidemic propagation. The overall conclusion is that epidemic differentiation is an emergent phenomenon dependent on temporal scale.
Significance. If the central claims hold after addressing methodological concerns, this work would be significant for the field of spatiotemporal epidemiology. It provides evidence that apparent differences in arboviral transmission are not intrinsic to the pathogens but arise from scale-dependent temporal dynamics, emphasizing the value of incorporating biological constraints and scale awareness in modeling. This could lead to improved understanding of urban epidemic dynamics and more effective intervention strategies. The use of large-scale georeferenced data and network reconstruction adds practical value, though robustness needs confirmation.
major comments (3)
- [Abstract] The abstract reports clear asymmetry between collapsed spatial and scale-dependent temporal parameters, yet provides no error bars, no explicit model equations, no data exclusion criteria, and no validation against held-out periods; post-hoc choice of the critical temporal window cannot be assessed.
- [Methods] Parameters are estimated by maximum likelihood on the same data used to declare scale dependence; the claim that unconstrained models capture only short-term co-occurrence while constrained ones reveal common structure is therefore partly tautological with the fitting procedure itself.
- [Results/Discussion] The central claim that differentiation is emergent at certain scales rests on the assumption that the pairwise interaction kernel with MLE and biological constraints correctly isolates the underlying transmission structure. Unmodeled factors such as vector density variation or reporting biases in the Recife data may confound the reported scale dependence, making it potentially an artifact of data collection rather than a true emergent property.
minor comments (2)
- [Abstract] Consider adding a brief mention of the specific biological constraints used in the model to enhance clarity for readers.
- [Figures] Ensure that any figures showing parameter estimates include error bars or confidence intervals for better interpretation of the collapse and scale dependence.
Simulated Author's Rebuttal
We thank the referee for their constructive and insightful comments, which have helped us identify areas for clarification and strengthening of the manuscript. We address each major comment point by point below, outlining our responses and planned revisions where appropriate.
read point-by-point responses
-
Referee: [Abstract] The abstract reports clear asymmetry between collapsed spatial and scale-dependent temporal parameters, yet provides no error bars, no explicit model equations, no data exclusion criteria, and no validation against held-out periods; post-hoc choice of the critical temporal window cannot be assessed.
Authors: We agree that the abstract, constrained by length, omits several technical details that would aid assessment. In the revised manuscript, we will expand the abstract to briefly reference the model equations (detailed in Methods), report error bars or confidence intervals on the key collapsed spatial and scale-dependent temporal parameters, and specify data exclusion criteria such as removal of cases lacking precise geolocation or with incomplete temporal records. For validation, we will note that the framework includes checks against held-out temporal periods (with results in supplementary materials). Regarding the critical temporal window, we will clarify its determination via systematic variation of window sizes and identification of the threshold where likelihood-ratio tests indicate statistically significant differentiation between diseases, rather than arbitrary post-hoc selection. These changes will be incorporated to improve transparency. revision: yes
-
Referee: [Methods] Parameters are estimated by maximum likelihood on the same data used to declare scale dependence; the claim that unconstrained models capture only short-term co-occurrence while constrained ones reveal common structure is therefore partly tautological with the fitting procedure itself.
Authors: We appreciate this point and will revise the Methods to eliminate any ambiguity. The biological constraints (e.g., minimum incubation and infectious periods drawn from independent virological literature) are imposed a priori and are not fitted from the Recife case data. The unconstrained model permits free estimation of all kernel parameters via MLE, while the constrained version enforces these external priors. Scale dependence is assessed by varying the temporal window size independently of the fitting step and observing the emergence of differentiation. We will add explicit text separating the sources of constraints from the data, include a sensitivity analysis demonstrating that the common underlying structure holds across reasonable constraint variations, and clarify that the short-term co-occurrence capture in unconstrained models is a direct consequence of allowing parameters to fit noise at small scales. This revision will strengthen the non-tautological nature of the comparison. revision: yes
-
Referee: [Results/Discussion] The central claim that differentiation is emergent at certain scales rests on the assumption that the pairwise interaction kernel with MLE and biological constraints correctly isolates the underlying transmission structure. Unmodeled factors such as vector density variation or reporting biases in the Recife data may confound the reported scale dependence, making it potentially an artifact of data collection rather than a true emergent property.
Authors: This concern is well-taken and highlights a genuine limitation in observational data. While the model infers pairwise interactions from georeferenced cases, factors like spatially varying vector density or differential reporting could influence patterns. However, the consistent collapse of spatial parameters across all three diseases argues against spatial confounders driving the results, as such biases would be expected to affect diseases similarly yet do not produce differentiation at small scales. Temporal scale dependence emerges robustly under constraints. In revision, we will add a dedicated limitations paragraph in the Discussion explicitly addressing these confounders, including how the large dataset size and focus on relative inter-disease differences provide some mitigation. We will also include supplementary robustness analyses, such as data subsampling to simulate reporting rate variations. We maintain that the emergent property interpretation is supported by the asymmetry and network reconstructions, but acknowledge that complete isolation of transmission dynamics would benefit from additional environmental covariates not present in the current dataset. revision: partial
Circularity Check
No significant circularity; derivation follows from MLE on external data
full rationale
The abstract describes a pairwise interaction kernel model whose parameters are estimated via maximum likelihood on the Recife georeferenced case data. Claims of scale-dependent temporal signatures, spatial parameter collapse, and differentiation between constrained versus unconstrained models are presented as outcomes of applying the fitted model to that data and inspecting the resulting networks. No equations, self-citations, or definitional steps are quoted that would reduce any central result to an input by construction. The distinction between short-term co-occurrence and common structure is an interpretive finding from the fits rather than a tautological renaming or self-referential constraint. The chain therefore remains self-contained against the external benchmark of the observed transmission events.
Axiom & Free-Parameter Ledger
free parameters (1)
- spatial and temporal kernel parameters
axioms (2)
- domain assumption Pairwise interactions between cases are sufficient to capture urban transmission dynamics
- domain assumption Biological constraints can be imposed by restricting the temporal kernel support
Forward citations
Cited by 1 Pith paper
-
Causal Edge Rees Algebras for Spatiotemporal Graphs
Causal Edge Rees Algebras encode the evolution of causal spatiotemporal graphs in a graded algebraic object, with a bridge detection theorem linking module dimensions to reductions in connected components.
Reference graph
Works this paper leans on
-
[1]
R.Markov Chains
Norris, J. R.Markov Chains. Cambridge Univer- sity Press, 1998
1998
-
[2]
A.; Peres, Y .; Wilmer, E
Levin, D. A.; Peres, Y .; Wilmer, E. L.Markov Chains and Mixing Times. American Mathemati- cal Society, 2017
2017
-
[3]
Physical Review E, 77(3), 036111, 2008
Estrada, E.; Hatano, N.Communicability in com- plex networks. Physical Review E, 77(3), 036111, 2008
2008
-
[4]
Ball, M. O. Computational complexity of network reliability analysis.IEEE Transactions on Relia- bility, 35(3), 230–239, 1986
1986
-
[5]
J.The Combinatorics of Network Re- liability
Colbourn, C. J.The Combinatorics of Network Re- liability. Oxford University Press, 1987
1987
-
[6]
Ricci curvature of Markov chains on metric spaces.Journal of Functional Analysis, 256(3), 810–864, 2009
Ollivier, Y . Ricci curvature of Markov chains on metric spaces.Journal of Functional Analysis, 256(3), 810–864, 2009
2009
-
[7]
Graph curvature for differentiating cancer net- works.Scientific Reports, 5, 12323, 2016
Sandhu, R.; Georgiou, T.; Reznik, E.; Zhu, L.; Kolesov, I.; Senbabaoglu, Y .; Tannenbaum, A. Graph curvature for differentiating cancer net- works.Scientific Reports, 5, 12323, 2016
2016
-
[8]
Ricci cur- vature of the Internet topology.IEEE INFOCOM, 2019
Ni, C.-C.; Lin, Y .-Y .; Luo, F.; Gao, J. Ricci cur- vature of the Internet topology.IEEE INFOCOM, 2019
2019
-
[9]
C.Socio-environmental determinants of dengue epidemics
Barcellos, C.; Sabroza, P. C.Socio-environmental determinants of dengue epidemics. Cadernos de Saúde Pública, 17, 77–86, 2001
2001
-
[10]
G.; Costa, M
Teixeira, M. G.; Costa, M. C. N.; Barreto, F.; Mota, E.Dengue: twenty-five years since its reemergence in Brazil. Cadernos de Saúde Pública, 25(Suppl 1), S7–S18, 2009
2009
-
[11]
A.; Nogueira, R
Honório, N. A.; Nogueira, R. M. R.; Codeço, C. T.; Carvalho, M. S.; Cruz, O. G.; Magalhães, M. A. F. M.; Lourenço-de-Oliveira, R.Spatial evalu- ation and modeling of dengue seroprevalence and vector density in Rio de Janeiro. PLoS Neglected Tropical Diseases, 3(11), e545, 2009
2009
-
[12]
J.; Rohani, P.Modeling Infectious Diseases in Humans and Animals
Keeling, M. J.; Rohani, P.Modeling Infectious Diseases in Humans and Animals. Princeton Uni- versity Press, 2008
2008
-
[13]
Topological persistence and simplification
Edelsbrunner, H.; Letscher, D.; Zomorodian, A. Topological persistence and simplification. In: Proceedings of the 41st Annual Symposium on Foundations of Computer Science (FOCS), 454– 463, 2000
2000
-
[14]
Computing persis- tent homology.Discrete and Computational Ge- ometry, 33(2), 249–274, 2005
Zomorodian, A.; Carlsson, G. Computing persis- tent homology.Discrete and Computational Ge- ometry, 33(2), 249–274, 2005
2005
-
[15]
Chazal, F.; Michel, B.; Rinaldo, A. An in- troduction to topological data analysis: funda- mental and practical aspects for data scientists. arXiv:1710.04019, 2016
-
[16]
Understanding climate variability using topological data analysis
Lorenz, D.; Junge, M.; Kramm, A. Understanding climate variability using topological data analysis. Chaos, 31(3), 033122, 2021
2021
-
[17]
A topo- logical representation of branching neuronal mor- phologies.Biosystems, 199, 104286, 2021
Kanari, L.; Dlotko, P.; Williams, S.; et al. A topo- logical representation of branching neuronal mor- phologies.Biosystems, 199, 104286, 2021
2021
-
[18]
Topological meth- ods in spatial epidemiology: detecting hotspots using persistent homology.Scientific Reports, 13, 2043, 2023
Bhatt, S.; Kumar, D.; Singh, A. Topological meth- ods in spatial epidemiology: detecting hotspots using persistent homology.Scientific Reports, 13, 2043, 2023
2043
-
[19]
S.; Souza-Santos, R.; Barcellos, C
Carvalho, M. S.; Souza-Santos, R.; Barcellos, C. Machine learning and spatial epidemiology: A new approach to understand urban arboviruses. Revista de Saúde Pública, 55, 44, 2021
2021
-
[20]
I.Satellite earth observa- tion data in epidemiological modeling of malaria, 9 dengue and West Nile virus: A scoping review
Parselia, E.; Kontoes, C.; Tsouni, A.; Had- jichristodoulou, C.; Kioutsioukis, I.; Magiorki- nis, G.; Stilianakis, N. I.Satellite earth observa- tion data in epidemiological modeling of malaria, 9 dengue and West Nile virus: A scoping review. Re- mote Sensing, 11(16), 1862, 2019
2019
-
[21]
The physics of communicability in complex net- works.Physics Reports, 514(3), 89–119, 2012
Estrada, E.; Hatano, N.; Benzi, M. The physics of communicability in complex net- works.Physics Reports, 514(3), 89–119, 2012. doi:10.1016/j.physrep.2012.01.006
-
[22]
Scientific Reports, 9(1), 16911, 2019
Massaro, E.; Kondor, D.; Ratti, C.Assessing the interplay between human mobility and mosquito- borne diseases in urban environments. Scientific Reports, 9(1), 16911, 2019
2019
-
[23]
I.; Simoy, M
Simoy, M. I.; Simoy, M. V .; Canziani, G. A.The effect of temperature on the population dynamics of Aedes aegypti. Ecological Modelling, 314, 100– 110, 2015
2015
-
[24]
Infectious Diseases of Poverty, 8, 1–12, 2019
Zheng, L.; Ren, H.-Y .; Shi, R.-H.; Lu, L.Spa- tiotemporal characteristics and primary influenc- ing factors of typical dengue fever epidemics in China. Infectious Diseases of Poverty, 8, 1–12, 2019
2019
-
[25]
C.; Brown, H
Moore, T. C.; Brown, H. E. EstimatingAedes ae- gyptiflight distance: meta-data analysis.Journal of Medical Entomology, 59(4), 1164–1170, 2022
2022
-
[26]
A.; et al
Pruszynski, C. A.; et al. Estimation of population age structure, daily survival rates, and potential to support dengue virus transmission for Florida KeysAedes aegypti.PLoS Neglected Tropical Dis- eases, 18(8), e0012350, 2024
2024
-
[27]
Geographical Analysis, 27(2), 93– 115, 1995
Anselin, L.Local Indicators of Spatial Associ- ation—LISA. Geographical Analysis, 27(2), 93– 115, 1995
1995
-
[28]
B.; Da Cunha, J
De Souza, D. B.; Da Cunha, J. T. S.; Dos Santos, E. F.; Correia, J. B.; Da Silva, H. P.; De Lima Filho, J. L.; Albuquerque, J.; San- tos, F. A. N. Using discrete Ricci curvatures to infer COVID-19 epidemic network fragility and systemic risk.Journal of Statistical Mechanics, 2021(5), 053501, 2021
2021
-
[29]
American Chemical So- ciety, 2012
Estrada, E.The Structure of Complex Networks: Theory and Applications. American Chemical So- ciety, 2012
2012
-
[30]
Forman–Ricci communicability curva- ture of graphs and networks.European Journal of Applied Mathematics, 1–25, 2025
Estrada, E. Forman–Ricci communicability curva- ture of graphs and networks.European Journal of Applied Mathematics, 1–25, 2025
2025
-
[31]
H.; De La Nuez, R.; Estrada, E
Grass-Boada, D. H.; De La Nuez, R.; Estrada, E. Graph/Network Reduction Based on Commu- nicability Vertex Similarity. 2025. (Manuscrito em processo de publicação.)
2025
-
[32]
P.Spatial Econometrics: From Cross- Sectional Data to Spatial Panels
Elhorst, J. P.Spatial Econometrics: From Cross- Sectional Data to Spatial Panels. Springer, 2014
2014
-
[33]
Wiley, 2015
Blangiardo, M.; Cameletti, M.Spatial and Spatio- Temporal Bayesian Models with R-INLA. Wiley, 2015
2015
-
[34]
Ferreira dos Santos, M.; Dos Santos Rodrigues de Melo, A.Hierarchical Bayesian Model- ing of Dengue in Recife, Brazil (2015–2024). arXiv:2510.13672, 2025
-
[35]
Zenodo.https://doi.org/10
Ferreira dos Santos, M.; Dos Santos Rodrigues de Melo, A.Spatial and Socioenvironmental Dengue Dataset of the Recife Metropolitan Area (2015–2024). Zenodo.https://doi.org/10. 5281/zenodo.17364863, 2025
2015
-
[36]
Zenodo.https://doi.org/10.5281/ zenodo.17849496, 2025
Ferreira dos Santos, M.; Dos Santos Rodrigues de Melo, A.Dengue Cases in Recife, Brazil (2015–2024): Clean and Geocoded Dataset (v1.0). Zenodo.https://doi.org/10.5281/ zenodo.17849496, 2025
2015
-
[37]
Ferreira dos Santos, M.Dengue Risk Model- ing and Network Curvature Analysis – Code Repository. GitHub. Disponível em:https: //github.com/DocDengueResearcher/ dengue-risk-recife-anonymous/blob/ main/Graph_Prototype_2_0.ipynb
-
[38]
T.; Ajelli, M.; et al.The effect of travel restrictions on COVID-19 spread
Chinazzi, M.; Davis, J. T.; Ajelli, M.; et al.The effect of travel restrictions on COVID-19 spread. Science, 368(6489), 395–400, 2020
2020
-
[39]
M.; Botai, J
Adeola, A. M.; Botai, J. O.; Olwoch, J. M.; Raut- enbach, C. J. H.; Adisa, O. M.Landsat-derived environmental metrics for mapping mosquito habi- tats. South African Geographical Journal, 99(1), 14–28, 2017
2017
-
[40]
Borges, I. V . G.; Musah, A.; Dutra, L. M. M.; et al.Interrelationship between precipitation and dengue in Recife. Frontiers in Public Health, 12, 1456043, 2024
2024
-
[41]
M.; et al.Dengue Monitoring Dashboard
Oliveira, M. M.; et al.Dengue Monitoring Dashboard. Cadernos de Saúde Pública, 38, e00252021, 2022. 10
2022
-
[42]
Santos, M. F., Melo, A. R. S., & Ricardo, C. L.Simulation SIR in Graphs: Het- erogeneous Curvature-Weighted Models. Zenodo (2025). Available at:https: //doi.org/10.5281/zenodo.17884569. doi:10.5281/zenodo.17884569
-
[43]
Santos, M. F.; Ricardo, C. L.; Melo, A. S. R.,Correlation-Weighted Communicability Cur- vature as a Structural Driver of Dengue Spread: A Bayesian Spatial Analysis of Recife (2015–2024), arXiv:2512.00315, 2025. Available at:https: //arxiv.org/abs/2512.00315
-
[44]
W.Systems of differential equations which are competitive or cooperative I: Limit sets
Hirsch, M. W.Systems of differential equations which are competitive or cooperative I: Limit sets. SIAM Journal on Mathematical Analysis, 16(2), 423–439, 1985
1985
-
[45]
L.Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems
Smith, H. L.Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems. Mathematical Surveys and Monographs, V ol. 41. American Mathematical So- ciety, 1995
1995
-
[46]
Kluwer Academic Publishers, 1988
Anselin, L.Spatial Econometrics: Methods and Models. Kluwer Academic Publishers, 1988
1988
-
[47]
P.; Pace, R
LeSage, J. P.; Pace, R. K.Introduction to Spatial Econometrics. CRC Press, 2009
2009
-
[48]
J.; Folmer, H.; Rey, S
Florax, R. J.; Folmer, H.; Rey, S. J. Specification searches in spatial econometrics. Regional Science and Urban Economics, 33(5), 557–579, 2003
2003
-
[49]
Approximate Bayesian inference for latent Gaussian models us- ing INLA
Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models us- ing INLA. Journal of the Royal Statistical Society B, 71(2), 319–392, 2009
2009
-
[50]
H.; Simpson, D.; Rue, H
Riebler, A.; Sørbye, S. H.; Simpson, D.; Rue, H. An intuitive Bayesian spatial model for dis- ease mapping that accounts for scaling. Statistical Methods in Medical Research, 25(4), 1145–1165, 2016
2016
-
[51]
An ex- plicit link between Gaussian fields and Gaussian Markov random fields: the SPDE approach
Lindgren, F.; Rue, H.; Lindström, J. An ex- plicit link between Gaussian fields and Gaussian Markov random fields: the SPDE approach. Jour- nal of the Royal Statistical Society B, 73(4), 423– 498, 2011. 11
2011
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.