Recognition: unknown
Logistic Gene Regulatory Networks: Prevention of Expression Shutdown, and Numerical Stability Beyond Hill Function
Pith reviewed 2026-05-09 14:34 UTC · model grok-4.3
The pith
Logistic functions fix the smoothness, numerical stability, and zero-basal-rate flaws of Hill functions in gene regulatory network models.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
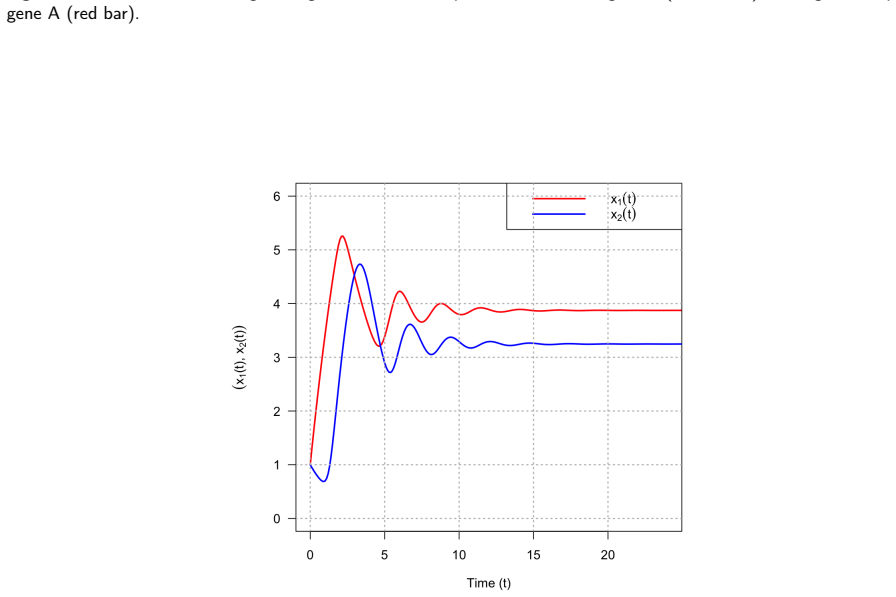
Logistic functions f±, defined to be globally C^∞, real for every real input, and strictly positive at zero, resolve the three structural flaws of Hill functions simultaneously. In a two-gene negative-feedback oscillator, the Routh-Hurwitz criterion shows local asymptotic stability for all positive parameter values and rules out Hopf bifurcation without time delays. For bistable positive autoregulation, saddle-node thresholds are given by explicit transcendental equations; with E. coli parameters the logistic basal rate produces off-state escape in approximately 44 minutes while the Hill model remains trapped indefinitely. The product-of-logistics De Morgan formalism translates Boolean logic
What carries the argument
The logistic activation functions f± that replace Hill functions, guaranteeing global smoothness, real arithmetic, and positive basal production rate.
Load-bearing premise
That logistic functions with positive basal production represent the underlying biology at least as accurately as Hill functions for the gene circuits and parameter ranges considered.
What would settle it
A laboratory measurement showing that a bistable positive-autoregulation circuit remains permanently off under conditions where the logistic model predicts escape within one hour, or a simulation run in which a Hill model with non-integer exponent produces no complex values or warnings.
Figures





read the original abstract
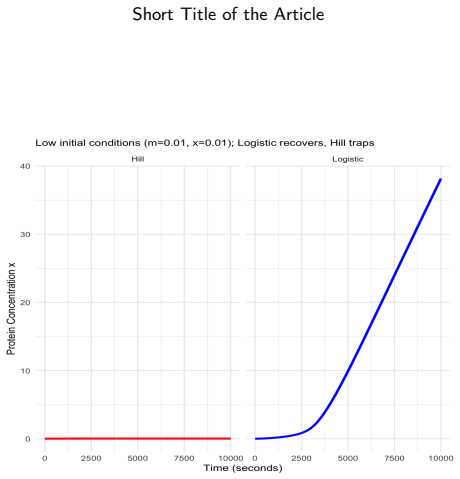
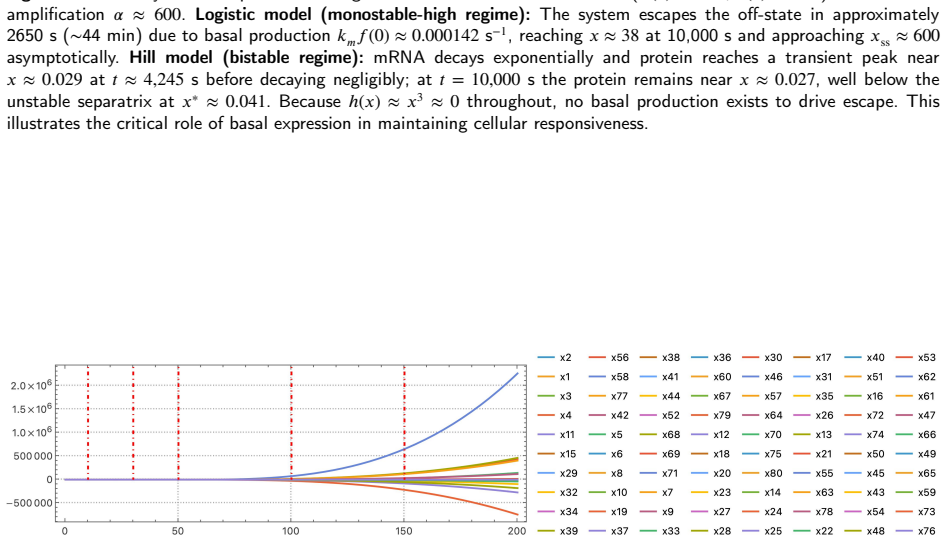
Hill functions, the standard tool for modelling gene regulatory networks, carry three structural flaws when the cooperativity exponent is non-integer: loss of global smoothness, silent complex-valued arithmetic corruption of ODE trajectories, and an identically zero basal production rate that traps bistable models in off-states. Logistic functions $f^\pm$, being globally $C^\infty$, real-valued for all arguments, and strictly positive at zero, resolve all three simultaneously. For a two-gene negative-feedback oscillator, local asymptotic stability is established for all positive parameters via the Routh--Hurwitz criterion, and no Hopf bifurcation is possible without time delays. For bistable positive autoregulation, saddle-node thresholds are characterised through explicit transcendental equations; with biophysically grounded \textit{E.~coli} parameters, basal logistic production drives off-state escape in $\approx 44$~min while the Hill model remains permanently trapped. The 11-gene Traynard cell-cycle Boolean network is translated automatically via the product-of-logistics De~Morgan formalism and integrated without warnings, all variables remaining bounded and non-negative. The De~Morgan framework places every repressor threshold at a positive measurable concentration, whereas the weighted-sum formulation of Samuilik et al.\ places repressor critical points at negative concentrations, rendering them biologically inert. On an 80-gene Boolean-derived ODE system with $n = 3.509$, the Hill solver entered silent complex-valued contamination at $t \approx 52.64$ and terminated near $t \approx 63$--$65$; the logistic formulation completed $t \in [0, 200]$ without a single warning. The always-positive production rate ensures full controllability, enabling sliding mode, model predictive, and feedback-linearisation strategies where Hill-based formulations fail.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes logistic functions f^± as replacements for Hill functions in gene regulatory network (GRN) models. It argues that logistics simultaneously resolve three Hill-function flaws for non-integer cooperativity: loss of global C^∞ smoothness, silent complex-valued arithmetic corruption of ODE trajectories, and identically zero basal production that traps bistable models in off-states. The paper establishes local asymptotic stability of a two-gene negative-feedback oscillator for all positive parameters via the Routh-Hurwitz criterion with no Hopf bifurcation possible without delays; derives saddle-node thresholds for bistable positive autoregulation via explicit transcendental equations; reports off-state escape in ≈44 min under biophysically grounded E. coli parameters; translates an 11-gene Traynard cell-cycle Boolean network via product-of-logistics De Morgan formalism; and demonstrates that an 80-gene Boolean-derived ODE system (n=3.509) completes t∈[0,200] without warnings while the Hill formulation enters complex-valued contamination at t≈52.64.
Significance. If the logistic basal rate is biologically appropriate, the work supplies a globally smooth, real-valued, and fully controllable framework for GRN modeling that avoids numerical artifacts and enables control-theoretic methods. The Routh-Hurwitz proof, explicit transcendental saddle-node conditions, and reproducible numerical demonstrations on both small and 80-gene systems constitute clear strengths. The De Morgan threshold placement at positive concentrations is a useful contrast to prior weighted-sum formulations. Significance is tempered by the need for stronger justification that non-zero basal production improves rather than distorts bistable dynamics.
major comments (2)
- [Bistable positive autoregulation] Bistable positive autoregulation section: the claim that logistic f^± 'prevents expression shutdown' rests on the finite mean escape time produced exactly by f(0)>0. The manuscript reports ≈44 min escape under 'biophysically grounded E. coli parameters' but does not list the explicit basal production rate, its literature source, or sensitivity analysis; without these, it is impossible to assess whether the result reflects measured promoter leakiness or post-hoc parameter selection that artifactually destabilizes the off-state.
- [Two-gene negative-feedback oscillator] Oscillator stability section: the Routh-Hurwitz criterion is applied to conclude local asymptotic stability for all positive parameters and no Hopf bifurcation without delays. The characteristic equation and the explicit Routh array entries should be provided so readers can verify the sign conditions independently; their absence makes the 'for all positive parameters' claim difficult to check.
minor comments (2)
- [Introduction / Methods] The explicit functional forms of the logistic activation f^+ and repression f^- (including the precise definition of the cooperativity parameter) are referenced but not displayed in the abstract or early sections; they should appear with equation numbers before any stability or bifurcation analysis.
- [Large-network numerical integration] In the 80-gene numerical example the cooperativity n=3.509 is stated without indicating whether it arises from a fit to data or is chosen for illustration; this detail affects reproducibility of the complex-value failure time t≈52.64.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed comments. We address each major point below and will revise the manuscript to incorporate the requested clarifications and supporting material.
read point-by-point responses
-
Referee: [Bistable positive autoregulation] Bistable positive autoregulation section: the claim that logistic f^± 'prevents expression shutdown' rests on the finite mean escape time produced exactly by f(0)>0. The manuscript reports ≈44 min escape under 'biophysically grounded E. coli parameters' but does not list the explicit basal production rate, its literature source, or sensitivity analysis; without these, it is impossible to assess whether the result reflects measured promoter leakiness or post-hoc parameter selection that artifactually destabilizes the off-state.
Authors: We agree that explicit documentation is required. In the revised manuscript we will state the precise basal production rate employed for the E. coli parameter set, cite the primary literature sources for measured promoter leakiness in E. coli, and add a sensitivity analysis that varies the basal rate over the range of experimentally reported values. This will show that off-state escape times remain on the order of tens of minutes for biophysically plausible leakiness levels. revision: yes
-
Referee: [Two-gene negative-feedback oscillator] Oscillator stability section: the Routh-Hurwitz criterion is applied to conclude local asymptotic stability for all positive parameters and no Hopf bifurcation without delays. The characteristic equation and the explicit Routh array entries should be provided so readers can verify the sign conditions independently; their absence makes the 'for all positive parameters' claim difficult to check.
Authors: We will include the full derivation in the revision. The updated section will present the Jacobian of the two-gene system, the resulting characteristic polynomial, and the complete Routh array with all entries and sign conditions. This will allow direct verification that the Hurwitz criteria are satisfied for every positive parameter combination. revision: yes
Circularity Check
No significant circularity; derivations are self-contained
full rationale
The paper derives local stability for the negative-feedback oscillator directly from the Routh-Hurwitz criterion applied to the logistic ODE system, obtains saddle-node thresholds for positive autoregulation via explicit transcendental equations, and performs direct numerical integration on the 11-gene and 80-gene networks. All results follow from the stated functional forms of f^±, the De Morgan translation rule, and externally supplied biophysical parameters without any fitting of outputs to inputs, self-referential definitions, or load-bearing self-citations. The contrast with Hill functions is presented as a transparent consequence of the basal-rate difference rather than a derived prediction.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math The logistic function is globally C^∞, real-valued for all real arguments, and strictly positive at zero
Reference graph
Works this paper leans on
-
[1]
An empirical extremum principle for the hill coefficient in ligand-protein interactions showing negative cooperativity
Abeliovich, H., 2005. An empirical extremum principle for the hill coefficient in ligand-protein interactions showing negative cooperativity. Biophysical journal 89, 76–79
2005
-
[2]
Atheoreticalexplorationofbirhythmicityinthep53-mdm2network
Abou-Jaoudé,W.,Chaves,M.,Gouzé,J.L.,2011. Atheoreticalexplorationofbirhythmicityinthep53-mdm2network. PLOSone6,e17075
2011
-
[3]
Enhancementofcellularmemorybyreducingstochastictransitions
Acar,M.,Becskei,A.,VanOudenaarden,A.,2005. Enhancementofcellularmemorybyreducingstochastictransitions. Nature435,228–232
2005
-
[4]
Quantitativemodelforgeneregulationbylambdaphagerepressor
Ackers,G.K.,Johnson,A.D.,Shea,M.A.,1982. Quantitativemodelforgeneregulationbylambdaphagerepressor. Proceedingsofthenational academy of sciences 79, 1129–1133
1982
-
[5]
The topology of the regulatory interactions predicts the expression pattern of the segment polarity genes in drosophila melanogaster
Albert, R., Othmer, H.G., 2003. The topology of the regulatory interactions predicts the expression pattern of the segment polarity genes in drosophila melanogaster. Journal of theoretical biology 223, 1–18
2003
-
[6]
An introduction to systems biology
Alon, U., 2007. An introduction to systems biology. Chapman & Hall/CRC, Boca Raton
2007
-
[7]
Cellular heterogeneity: do differences make a difference? Cell 141, 559–563
Altschuler, S.J., Wu, L.F., 2010. Cellular heterogeneity: do differences make a difference? Cell 141, 559–563
2010
-
[8]
Engineering stability in gene networks by autoregulation
Becskei, A., Serrano, L., 2000. Engineering stability in gene networks by autoregulation. Nature 405, 590–593
2000
-
[9]
Exploring Logistic Functions as Robust Alternatives to Hill Functions in Genetic Network Modeling
Belgacem, I., 2025. Exploring logistic functions as robust alternatives to hill functions in genetic network modeling. arXiv preprint arXiv:2512.14325
work page internal anchor Pith review Pith/arXiv arXiv 2025
-
[10]
Theprobabilisticconvolutionregularizationofzenohybridsystems,in:201918th European Control Conference (ECC), IEEE
Belgacem,I.,Bensalah,H.,Cherki,B.,Edwards,R.,2019. Theprobabilisticconvolutionregularizationofzenohybridsystems,in:201918th European Control Conference (ECC), IEEE. pp. 750–757
2019
-
[11]
Reduction and stability analysis of a transcription–translation model of rna polymerase
Belgacem, I., Casagranda, S., Grac, E., Ropers, D., Gouzé, J.L., 2018. Reduction and stability analysis of a transcription–translation model of rna polymerase. Bulletin of Mathematical Biology 80, 294–318
2018
-
[12]
Belgacem,I.,Edwards,R.,Farcot,E.,2025. Computer-aidedanalysisofhigh-dimensionalGlassnetworks:periodicity,chaos,andbifurcations in a ring circuit. Chaos: An Interdisciplinary Journal of Nonlinear Science doi:10.1063/5.0243955
-
[13]
Belgacem, I., Gouzé, J.L., 2012. Global stability of full open reversible Michaelis–Menten reactions, in: 8th IFAC Symposium on Advanced Control of Chemical Processes, Singapore. pp. 591–596. doi:10.3182/20120710-4-SG-2026.00039
-
[14]
Analysis and reduction of transcription translation coupled models for gene expression
Belgacem, I., Gouzé, J.L., 2013a. Analysis and reduction of transcription translation coupled models for gene expression. IFAC Proceedings Volumes 46, 36–41
-
[15]
Global stability of enzymatic chains of full reversible michaelis-menten reactions
Belgacem, I., Gouzé, J.L., 2013b. Global stability of enzymatic chains of full reversible michaelis-menten reactions. Acta biotheoretica 61, 425–436
-
[16]
Belgacem,I.,Gouzé,J.L.,2013. Stabilityanalysisandreductionofgenetranscriptionmodels,in:IEEE52ndAnnualConferenceonDecision and Control (CDC’13), Florence, Italy. pp. 2691–2696. doi:10.1109/CDC.2013.6760289
-
[17]
Belgacem, I., Gouzé, J.L., 2014. Mathematical study of the global dynamics of a concave gene expression model, in: 22nd Mediterranean Conference on Control and Automation (MED’14), Palermo, Italy. pp. 1341–1346. doi:10.1109/MED.2014.6961562
-
[18]
Control of negative feedback loops in genetic networks, in: 2020 59th IEEE Conference on Decision and Control (CDC), IEEE
Belgacem, I., Gouzé, J.L., Edwards, R., 2020. Control of negative feedback loops in genetic networks, in: 2020 59th IEEE Conference on Decision and Control (CDC), IEEE. pp. 5098–5105
2020
-
[19]
Belgacem, I., Gouzé, J.L., Edwards, R., 2021. Control of negative feedback loops in genetic networks, in: Proceedings of the 59th IEEE Conference on Decision and Control (CDC), Jeju Island, Republic of Korea. doi:10.1109/CDC42340.2020.9304088
-
[20]
Belgacem, I., Grac, E., Ropers, D., Gouzé, J.L., 2014. Stability analysis of a reduced transcription-translation model of RNA polymerase, in: IEEE 53rd Conference on Decision and Control (CDC), Los Angeles, California, USA. pp. 3924–3929. doi:10.1109/CDC.2014.7039999
-
[21]
Quantification of protein half-lives in the budding yeast proteome
Belle, A., Tanay, A., Bitincka, L., Shamir, R., O’Shea, E.K., 2006. Quantification of protein half-lives in the budding yeast proteome. Proceedings of the National Academy of Sciences 103, 13004–13009
2006
-
[22]
Modeling and analysis of gene regulatory networks, in: Modeling in computational biology and biomedicine: A multidisciplinary endeavor
Bernot, G., Comet, J.P., Richard, A., Chaves, M., Gouzé, J.L., Dayan, F., 2012. Modeling and analysis of gene regulatory networks, in: Modeling in computational biology and biomedicine: A multidisciplinary endeavor. Springer, pp. 47–80
2012
-
[23]
Global analysis of mrna decay and abundance in escherichia coliatsingle-generesolutionusingtwo-colorfluorescentdnamicroarrays
Bernstein, J.A., Khodursky, A.B., Lin, P.H., Lin-Chao, S., Cohen, S.N., 2002. Global analysis of mrna decay and abundance in escherichia coliatsingle-generesolutionusingtwo-colorfluorescentdnamicroarrays. ProceedingsoftheNationalAcademyofSciences99,9697–9702. First Author et al.: Page 35 of 37 Short Title of the Article
2002
-
[24]
Causal reasoning on boolean control networks based on abduction: theory and application to cancer drug discovery
Biane, C., Delaplace, F., 2018. Causal reasoning on boolean control networks based on abduction: theory and application to cancer drug discovery. IEEE/ACM transactions on computational biology and bioinformatics 16, 1574–1585
2018
-
[25]
Transcriptionalregulationbythenumbers:models
Bintu,L.,Buchler,N.E.,Garcia,H.G.,Gerland,U.,Hwa,T.,Kondev,J.,Phillips,R.,2005. Transcriptionalregulationbythenumbers:models. Current opinion in genetics & development 15, 116–124
2005
-
[26]
Hill function-based models of transcriptional switches: impact of specific, nonspecific, functional and nonfunctional binding
Bottani, S., Veitia, R.A., 2017. Hill function-based models of transcriptional switches: impact of specific, nonspecific, functional and nonfunctional binding. Biological Reviews 92, 953–963
2017
-
[27]
Qualitativecontrolofundesiredoscillationsinageneticnegativefeedbackloopwithuncertain measurements
Chambon,L.,Belgacem,I.,Gouzé,J.L.,2020. Qualitativecontrolofundesiredoscillationsinageneticnegativefeedbackloopwithuncertain measurements. Automatica 112, 108642
2020
-
[28]
The properties of logistic function and applications to neural network approximation
Chen, Z., Cao, F., 2013. The properties of logistic function and applications to neural network approximation. Journal of Computational Analysis and Applications 15, 1046–1056
2013
-
[29]
How to make a biological switch
Cherry, J.L., Adler, F.R., 2000. How to make a biological switch. Journal of theoretical biology 203, 117–133
2000
-
[30]
Mathematical modelling of molecular pathways enabling tumour cell invasion and migration
Cohen, D.P.A., Martignetti, L., Robine, S., Barillot, E., Zinovyev, A., Calzone, L., 2015. Mathematical modelling of molecular pathways enabling tumour cell invasion and migration. PLoS computational biology 11, e1004571. doi:10.1371/journal.pcbi.1004571
-
[31]
Biomolecular feedback systems
Del Vecchio, D., Murray, R.M., 2015. Biomolecular feedback systems. Princeton University Press Princeton, NJ
2015
-
[32]
A synthetic oscillatory network of transcriptional regulators
Elowitz, M.B., Leibler, S., 2000. A synthetic oscillatory network of transcriptional regulators. Nature 403, 335
2000
-
[33]
Enciso, J., Mayani, H., Mendoza, L., Pelayo, R., 2016. Modeling the pro-inflammatory tumor microenvironment in acute lymphoblastic leukemia predicts a breakdown of hematopoietic-mesenchymal communication networks. Frontiers in physiology 7, 349. doi:10.3389/ fphys.2016.00349
-
[34]
Biochemistry 39, 11074–11083
Falcon,C.M.,Matthews,K.S.,2000.Operatordnasequencevariationenhanceshighaffinitybindingbyhingehelixmutantsoflactoserepressor protein. Biochemistry 39, 11074–11083
2000
-
[35]
A computational framework for optimal and model predictive control of stochastic gene regulatory networks
Faquir, H., Pájaro, M., Otero-Muras, I., 2025. A computational framework for optimal and model predictive control of stochastic gene regulatory networks. IEEE Transactions on Computational Biology and Bioinformatics
2025
-
[36]
Chaosinaringcircuit
Farcot,E.,Best,S.,Edwards,R.,Belgacem,I.,Xu,X.,Gill,P.,2019. Chaosinaringcircuit. Chaos:AnInterdisciplinaryJournalofNonlinear Science 29, 043103
2019
-
[37]
An introduction to probability theory and its applications
Feller, W., et al., 1971. An introduction to probability theory and its applications. Wiley New York
1971
-
[38]
Construction of a genetic toggle switch in escherichia coli
Gardner, T.S., Cantor, C.R., Collins, J.J., 2000. Construction of a genetic toggle switch in escherichia coli. Nature 403, 339–342
2000
-
[39]
On sharpness of error bounds for univariate approximation by single hidden layer feedforward neural networks
Goebbels, S., 2020. On sharpness of error bounds for univariate approximation by single hidden layer feedforward neural networks. Results in Mathematics 75, 109
2020
-
[40]
Thefive-parameterlogistic:acharacterizationandcomparisonwiththefour-parameterlogistic
Gottschalk,P.G.,Dunn,J.R.,2005. Thefive-parameterlogistic:acharacterizationandcomparisonwiththefour-parameterlogistic. Analytical biochemistry 343, 54–65
2005
-
[41]
Solving ordinary differential equations I: Nonstiff problems
Hairer, E., Wanner, G., Nørsett, S.P., 1993. Solving ordinary differential equations I: Nonstiff problems. Springer
1993
-
[42]
Corrected hill function in stochastic gene regulatory networks
Hernández-García, M.E., Velázquez-Castro, J., 2023. Corrected hill function in stochastic gene regulatory networks. arXiv preprint arXiv:2307.03057
-
[43]
Multiple regulation of the galactose operon—genetic evidence for a distinct site in the galactose operon that responds to capr gene regulation in escherichia coli k-12
Hua, S.S., Markovitz, A., 1974. Multiple regulation of the galactose operon—genetic evidence for a distinct site in the galactose operon that responds to capr gene regulation in escherichia coli k-12. Proceedings of the National Academy of Sciences 71, 507–511. doi:10.1073/ pnas.71.2.507
1974
-
[44]
Effects of promoter leakage on dynamics of gene expression
Huang, L., Yuan, Z., Liu, P., Zhou, T., 2015. Effects of promoter leakage on dynamics of gene expression. BMC systems biology 9, 16. doi:10.1186/s12918-015-0157-z
-
[45]
Mathematical modeling in systems biology: an introduction
Ingalls, B.P., 2013. Mathematical modeling in systems biology: an introduction. MIT press
2013
-
[46]
Genetic regulatory mechanisms in the synthesis of proteins
Jacob, F., Monod, J., 1961. Genetic regulatory mechanisms in the synthesis of proteins. Journal of molecular biology 3, 318–356
1961
-
[47]
Basal leakage in oscillation: Coupled transcriptional and translational control using feed-forward loops
Joanito, I., Yan, C.C.S., Chu, J.W., Wu, S.H., Hsu, C.P., 2020. Basal leakage in oscillation: Coupled transcriptional and translational control using feed-forward loops. PLOS Computational Biology 16, e1007740
2020
-
[48]
Metabolic stability and epigenesis in randomly constructed genetic nets
Kauffman, S.A., 1969. Metabolic stability and epigenesis in randomly constructed genetic nets. Journal of theoretical biology 22, 437–467
1969
-
[49]
Stochastic gene expression modeling with hill function for switch-like gene responses
Kim, H., Gelenbe, E., 2011. Stochastic gene expression modeling with hill function for switch-like gene responses. IEEE/ACM Transactions on Computational Biology and Bioinformatics 9, 973–979
2011
-
[50]
Distributionofcellsizeingrowingculturesofbacteriaandtheapplicabilityofthecollins-richmondprinciple
Koch,A.,1966. Distributionofcellsizeingrowingculturesofbacteriaandtheapplicabilityofthecollins-richmondprinciple. Microbiology 45, 409–417
1966
-
[51]
Models of genetic networks with given properties
Kozlovska, O., Sadyrbaev, F., 2022. Models of genetic networks with given properties. WSEAS Transactions on Computer Research 10, 43–49
2022
-
[52]
In search of chaos in genetic systems
Kozlovska, O., Sadyrbaev, F., 2024. In search of chaos in genetic systems. Chaos Theory and Applications 6, 13–18
2024
-
[53]
Modeling networks of four elements
Kozlovska, O., Sadyrbaev, F., 2025. Modeling networks of four elements. Computation 13, 123
2025
-
[54]
Diya–auniversal light illumination platform for multiwell plate cultures
Kumar,S.,Anastassov,S.,Aoki,S.K.,Falkenstein,J.,Chang,C.H.,Frei,T.,Buchmann,P.,Argast,P.,Khammash,M.,2023. Diya–auniversal light illumination platform for multiwell plate cultures. Iscience 26
2023
-
[55]
Sigmoid functions: some approximation and modelling aspects
Kyurkchiev, N., Markov, S., 2015. Sigmoid functions: some approximation and modelling aspects. LAP LAMBERT Academic Publishing, Saarbrucken 4, 34
2015
-
[56]
Quantifyingabsoluteproteinsynthesisratesrevealsprinciplesunderlyingallocation of cellular resources
Li,G.W.,Burkhardt,D.,Gross,C.,Weissman,J.S.,2014. Quantifyingabsoluteproteinsynthesisratesrevealsprinciplesunderlyingallocation of cellular resources. Cell 157, 624–635
2014
-
[57]
Genetic toggle switch without cooperative binding
Lipshtat, A., Loinger, A., Balaban, N.Q., Biham, O., 2006. Genetic toggle switch without cooperative binding. Physical review letters 96, 188101
2006
-
[58]
Deep model predictive control of gene expression in thousands of single cells
Lugagne, J.B., Blassick, C.M., Dunlop, M.J., 2024. Deep model predictive control of gene expression in thousands of single cells. Nature Communications 15, 2148
2024
-
[59]
Negative auto-regulation increases the input dynamic-range of the arabinose system of escherichia coli
Madar, D., Dekel, E., Bren, A., Alon, U., 2011. Negative auto-regulation increases the input dynamic-range of the arabinose system of escherichia coli. BMC systems biology 5, 111. First Author et al.: Page 36 of 37 Short Title of the Article
2011
-
[60]
Quantitativemodelingoftranscription andtranslationofan all-e.colicell-freesystem
Marshall,R., Noireaux,V.,2019. Quantitativemodelingoftranscription andtranslationofan all-e.colicell-freesystem. Scientificreports9, 11980
2019
-
[61]
Precise modulation of transcription factor levels identifies features underlying dosage sensitivity
Naqvi,S.,Kim,S.,Hoskens,H.,Matthews,H.S.,Spritz,R.A.,Klein,O.D.,Hallgrímsson,B.,Swigut,T.,Claes,P.,Pritchard,J.K.,etal.,2023. Precise modulation of transcription factor levels identifies features underlying dosage sensitivity. Nature genetics 55, 841–851
2023
-
[62]
Protein degradation in escherichia coli: I
Nath, K., Koch, A.L., 1970. Protein degradation in escherichia coli: I. measurement of rapidly and slowly decaying components. Journal of Biological Chemistry 245, 2889–2900
1970
-
[63]
Genetic circuit design automation
Nielsen, A.A., Der, B.S., Shin, J., Vaidyanathan, P., Paralanov, V., Strychalski, E.A., Ross, D., Densmore, D., Voigt, C.A., 2016. Genetic circuit design automation. Science 352, aac7341
2016
-
[64]
Quality and position of the three lac operators of e
Oehler, S., Amouyal, M., Kolkhof, P., von Wilcken-Bergmann, B., Müller-Hill, B., 1994. Quality and position of the three lac operators of e. coli define efficiency of repression. The EMBO journal 13, 3348–3355
1994
-
[65]
Multistability in the lactose utilization network of escherichia coli
Ozbudak, E.M., Thattai, M., Lim, H.N., Shraiman, B.I., Van Oudenaarden, A., 2004. Multistability in the lactose utilization network of escherichia coli. Nature 427, 737–740
2004
-
[66]
Comparing different ode modelling approaches for gene regulatory networks
Polynikis, A., Hogan, S., di Bernardo, M., 2009. Comparing different ode modelling approaches for gene regulatory networks. Journal of theoretical biology 261, 511–530
2009
-
[67]
Tuning transcriptional regulation through signaling: a predictive theory of allosteric induction
Razo-Mejia, M., Barnes, S.L., Belliveau, N.M., Chure, G., Einav, T., Lewis, M., Phillips, R., 2018. Tuning transcriptional regulation through signaling: a predictive theory of allosteric induction. Cell systems 6, 456–469
2018
-
[68]
Pharmacodynamic models: parameterizing the hill equation, michaelis-menten, the logistic curve, and relationships among these models
Reeve, R., Turner, J.R., 2013. Pharmacodynamic models: parameterizing the hill equation, michaelis-menten, the logistic curve, and relationships among these models. Journal of biopharmaceutical statistics 23, 648–661
2013
-
[69]
On modelling of genetic regulatory networks
Sadyrbaev, F., Samuilik, I., Sengileyev, V., 2021. On modelling of genetic regulatory networks. WSEAS Transactions on Electronics 12, 73
2021
-
[70]
On coexistence of inhibition and activation in genetic regulatory networks, in: International Conference on Numerical Analysis and Applied Mathematics 2021, ICNAAM 2021, AIP PRESS
Sadyrbaev, F., Sengileyev, V., Silvans, A., 2023. On coexistence of inhibition and activation in genetic regulatory networks, in: International Conference on Numerical Analysis and Applied Mathematics 2021, ICNAAM 2021, AIP PRESS
2023
-
[71]
BMC systems biology 3, 1
Sahin,Ö.,Fröhlich,H.,Löbke,C.,Korf,U.,Burmester,S.,Majety,M.,Mattern,J.,Schupp,I.,Chaouiya,C.,Thieffry,D.,etal.,2009.Modeling erbb receptor-regulated g1/s transition to find novel targets for de novo trastuzumab resistance. BMC systems biology 3, 1
2009
-
[72]
Genetic engineering–construction of a network of arbitrary dimension with periodic attractor
Samuilik, I., Sadyrbaev, F., 2022. Genetic engineering–construction of a network of arbitrary dimension with periodic attractor. Vibroengi- neering Procedia 46, 67–72
2022
-
[73]
Mathematicalmodelingofthree-dimensionalgeneticregulatorynetworksusinglogisticand gompertz functions
Samuilik,I.,Sadyrbaev,F.,Ogorelova,D.,2022. Mathematicalmodelingofthree-dimensionalgeneticregulatorynetworksusinglogisticand gompertz functions. WSEAS Transactions on systems and control 17, 101107
2022
-
[74]
Ontheuseofthehillfunctionsinmathematicalmodelsofgeneregulatorynetworks
Santillán,M.,2008. Ontheuseofthehillfunctionsinmathematicalmodelsofgeneregulatorynetworks. MathematicalModellingofNatural Phenomena 3, 85–97
2008
-
[75]
Sawlekar, R., Montefusco, F., Kulkarni, V., Bates, D.G., 2015. Biomolecular implementation of a quasi sliding mode feedback controller basedondnastranddisplacementreactions,in:201537thAnnualInternationalConferenceoftheIEEEEngineeringinMedicineandBiology Society (EMBC), IEEE. pp. 949–952
2015
-
[76]
Detailed map of a cis-regulatory input function
Setty, Y., Mayo, A.E., Surette, M.G., Alon, U., 2003. Detailed map of a cis-regulatory input function. Proceedings of the National Academy of Sciences 100, 7702–7707
2003
-
[77]
Revealing gene regulation-based neural network computing in bacteria
Somathilaka, S.S., Balasubramaniam, S., Martins, D.P., Li, X., 2023. Revealing gene regulation-based neural network computing in bacteria. Biophysical Reports 3
2023
-
[78]
Traynard, P., Fauré, A., Fages, F., Thieffry, D., 2016. Logical model specification aided by model-checking techniques: application to the mammalian cell cycle regulation. Bioinformatics (Oxford, England) 32, i772–i780. doi:10.1093/bioinformatics/btw457
-
[79]
Verlingue, L., Dugourd, A., Stoll, G., Barillot, E., Calzone, L., Londoño-Vallejo, A., 2016. A comprehensive approach to the molecular determinants of lifespan using a boolean model of geroconversion. Aging cell 15, 1018–1026. doi:10.1111/acel.12504
-
[80]
The galactose regulon of escherichia coli
Weickert, M.J., Adhya, S., 1993. The galactose regulon of escherichia coli. Molecular microbiology 10, 245–251
1993
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.