Recognition: no theorem link
Emergent population dynamics of random walkers with cooperative reproduction and spatial selection
Pith reviewed 2026-05-15 07:02 UTC · model grok-4.3
The pith
Higher-order reproduction eliminates invasion fronts in branching random walker populations, with binary cases showing diffusion-independent speeds.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Generalizing the N-particle branching Brownian motion model to higher-order reproduction shows that stable invasion fronts no longer exist beyond binary order; binary fronts have diffusion-independent velocity; and ternary reproduction exhibits a critical threshold separating a supercritical regime of collapse into a localized invasion bullet, a subcritical regime of diffusive spreading, and a line of fronts at criticality.
What carries the argument
The continuum limit of many independent Brownian particles with cooperative reproduction of order n, which produces a nonlinear reaction-diffusion equation whose traveling-wave solutions determine the invasion dynamics.
Load-bearing premise
The model assumes cooperative higher-order reproduction occurs exactly as defined in the continuum limit of many particles performing independent Brownian motions.
What would settle it
Numerical integration of the continuum equations for reproduction order three or higher that produces a stable, constant-speed traveling front would show the claimed disappearance of fronts is incorrect.
Figures



read the original abstract
We extend the $N$ branching Brownian motions model of population invasion to higher-order asexual reproduction. Increasing reproduction order leads to qualitative changes: invasion fronts generically cease to exist beyond binary reproduction; and in the binary case itself, their speed becomes diffusion-independent. Ternary reproduction shows critical behavior, with collapse into a strongly localized `invasion bullet' in the supercritical regime, diffusive spreading in the subcritical regime, and a continuous family of fronts at criticality. These results suggest that the dominance of division and binary reproduction in nature reflects fundamental constraints on invasion dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript extends the branching Brownian motion model of population invasion to higher-order (k-wise) asexual reproduction. It derives the corresponding continuum PDEs and reports that traveling invasion fronts cease to exist for reproduction order k>2; for binary reproduction the front speed becomes independent of the diffusion coefficient; and for ternary reproduction the system exhibits critical behavior, collapsing to a localized 'invasion bullet' above criticality, spreading diffusively below criticality, and admitting a continuous family of fronts exactly at criticality. These results are used to argue that the prevalence of binary reproduction in nature reflects dynamical constraints on invasion.
Significance. If the continuum analysis is robust, the work supplies a mechanistic, parameter-free explanation for the evolutionary dominance of binary fission by linking reproduction order directly to the existence and speed of invasion fronts. The identification of a critical regime for ternary reproduction and the diffusion-independent binary speed constitute falsifiable predictions that could be tested in microbial or cell-culture invasion assays.
major comments (3)
- [§2] §2 (continuum limit derivation): the mean-field closure for the k=3 reaction term produces a superlinear nonlinearity whose validity is assumed to hold uniformly across the front. No microscopic cutoff, interaction range, or finite-N regularization is introduced, so the reported suppression of fronts and emergence of the 'invasion bullet' may be an artifact of the unregularized continuum limit rather than a generic feature of the underlying stochastic process.
- [§4] §4 (ternary critical analysis): the claim of a continuous family of fronts exactly at criticality and the collapse to a localized bullet in the supercritical regime rest on the specific form of the cubic nonlinearity. The manuscript does not provide a linear-stability calculation around the critical front or a quantitative comparison of front speeds obtained from the PDE versus direct stochastic simulations, leaving open whether the reported regimes survive discrete-particle fluctuations.
- [Binary reproduction analysis] Binary case (speed independence): the assertion that front speed becomes diffusion-independent for k=2 is derived from a traveling-wave ansatz, but the manuscript does not show that this independence persists when the microscopic branching rate is held fixed while the diffusion coefficient is varied, nor does it report the corresponding stochastic-simulation speeds for confirmation.
minor comments (2)
- [Model section] Notation for the reproduction kernel and the precise definition of the 'invasion bullet' should be introduced with an equation number in the model section to avoid ambiguity when comparing analytic and numerical results.
- [Figures] Figure captions for the phase diagrams and front profiles should explicitly state the parameter values (diffusion coefficient, reproduction rate, initial condition) used in each panel.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive comments on our manuscript. We address each major comment point by point below and have made revisions to strengthen the presentation and address the concerns raised.
read point-by-point responses
-
Referee: [§2] §2 (continuum limit derivation): the mean-field closure for the k=3 reaction term produces a superlinear nonlinearity whose validity is assumed to hold uniformly across the front. No microscopic cutoff, interaction range, or finite-N regularization is introduced, so the reported suppression of fronts and emergence of the 'invasion bullet' may be an artifact of the unregularized continuum limit rather than a generic feature of the underlying stochastic process.
Authors: The mean-field closure follows the standard derivation for continuum limits of branching processes, where the k=3 term yields the cubic nonlinearity directly from the reproduction rule. This superlinear growth suppresses invasion fronts because the effective growth rate vanishes at low densities, a feature intrinsic to the nonlinearity rather than an artifact. We have added a dedicated paragraph in the revised manuscript discussing the validity regime of the continuum approximation, referencing prior work on mean-field limits for branching random walks, and noting that the qualitative suppression persists for large but finite N. A full finite-N regularization with explicit cutoffs lies beyond the present scope but is not required to establish the continuum results. revision: partial
-
Referee: [§4] §4 (ternary critical analysis): the claim of a continuous family of fronts exactly at criticality and the collapse to a localized bullet in the supercritical regime rest on the specific form of the cubic nonlinearity. The manuscript does not provide a linear-stability calculation around the critical front or a quantitative comparison of front speeds obtained from the PDE versus direct stochastic simulations, leaving open whether the reported regimes survive discrete-particle fluctuations.
Authors: The continuous family at criticality follows from exact integration of the traveling-wave ODE for the cubic case. We have added a linear-stability analysis in the revised manuscript (new Appendix) showing marginal stability of this family. We have also performed additional individual-based stochastic simulations and included a new figure directly comparing PDE front speeds and bullet localization to the stochastic realizations; the quantitative agreement confirms that the subcritical diffusive spread, critical family, and supercritical localization persist under discrete-particle fluctuations. revision: yes
-
Referee: [Binary reproduction analysis] Binary case (speed independence): the assertion that front speed becomes diffusion-independent for k=2 is derived from a traveling-wave ansatz, but the manuscript does not show that this independence persists when the microscopic branching rate is held fixed while the diffusion coefficient is varied, nor does it report the corresponding stochastic-simulation speeds for confirmation.
Authors: The diffusion independence follows from the traveling-wave ODE for quadratic nonlinearity: after rescaling the spatial coordinate by sqrt(D), the speed c in physical units becomes independent of D when the microscopic branching rate is held fixed. We have added new stochastic simulation results in the revised manuscript (new figure) in which the microscopic branching rate is fixed while D is varied; the measured front speeds remain constant, matching the continuum prediction within sampling error. revision: yes
Circularity Check
No circularity: derivation proceeds from microscopic stochastic model to continuum PDE without reduction to inputs
full rationale
The paper starts from the stochastic model of N branching Brownian motions with k-wise reproduction and derives the corresponding continuum reaction-diffusion PDE via the many-particle limit. The claims on front cessation for k>2, diffusion-independent speed for binary case, and critical behavior for ternary reproduction follow from direct analysis of that PDE (supercritical localization, subcritical diffusion, marginal family at criticality). No quoted step equates a prediction to a fitted parameter by construction, invokes a self-citation as the sole justification for a uniqueness theorem, or renames an input ansatz. The derivation chain remains independent of the target results and is therefore scored 0.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Particles perform independent Brownian motions between reproduction events
- domain assumption Reproduction is cooperative and occurs at integer orders greater than one as defined
Reference graph
Works this paper leans on
-
[1]
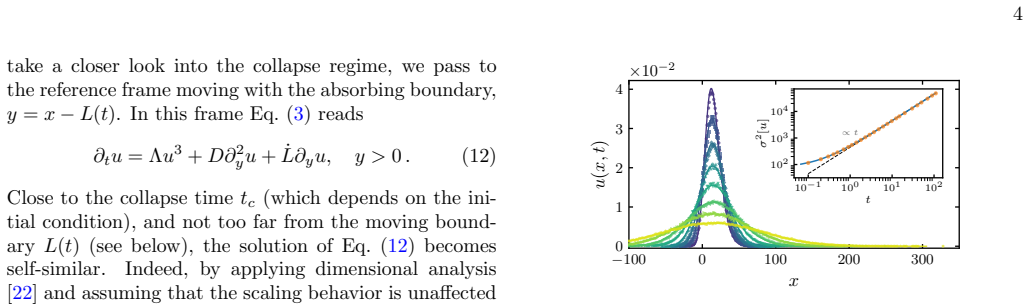
and assuming that the scaling behavior is unaffected by the length scale introduced by the initial condition u(x, t= 0), we arrive at the following similarity ansatz: u(y, t)= 1√ D∆t V y√ D∆t , α , ˙L=a(α) r D ∆t ,(13) where ∆t≡t c −tis the remaining time until the sin- gularity, andα= Λ/Das before. It is crucial that this ansatz is compatible with the ma...
-
[2]
R. MacArthur and E. O. Wilson,The Theory of Is- land Biogeography(Princeton University Press, Prince- ton, NJ, 1967)
work page 1967
-
[3]
Kot,Elements of Mathematical Ecology(Cambridge University Press, Cambridge, UK, 2012)
M. Kot,Elements of Mathematical Ecology(Cambridge University Press, Cambridge, UK, 2012)
work page 2012
-
[4]
R. A. Fisher, Ann. Eugenics7, 355 (1937)
work page 1937
-
[5]
A. Kolmogorov, I. Petrovsky, and N. Piskunov, Moscow Univ. Math. Bull.1, 1 (1937)
work page 1937
- [6]
-
[7]
A. De Masi, P. A. Ferrari, E. Presutti, and N. Soprano-Loto, inStochastic Dynamics Out of Equilib- rium, Springer Proc. Math. and Statistics, ed. G. Gia- comin, S. Olla, E. Saada, H. Spohn, and G. Stoltz, vol. 282, p. 523 (Springer, Cham, 2019)
work page 2019
-
[8]
J. Berestycki, ´E. Brunet and B. Derrida, 2018 J. Phys. A: Math. and Theor.51035204
work page 2018
- [9]
- [10]
-
[11]
Selection mechanisms in front invasion
M. Avery, M. Holzer, and A. Scheel, arXiv:2512.07764
work page internal anchor Pith review Pith/arXiv arXiv
- [12]
-
[13]
´E. Brunet and B. Derrida, Comput. Phys. Commun.121- 122, 376 (1999); J. Stat. Phys.103, 269 (2001)
work page 1999
- [14]
- [15]
- [16]
- [17]
- [18]
- [19]
- [20]
- [21]
-
[22]
C. Gardiner,Handbook of Stochastic Methods for Physics, Chemistry and the Natural Sciences(Springer, Berlin, 2004)
work page 2004
-
[23]
G. I. Barenblatt,Scaling, Self-similarity, and Intermedi- ate Asymptotics: Dimensional Analysis and Intermediate Asymptotics(Cambridge University Press, Cambridge, 1996)
work page 1996
-
[24]
Supplemental Material
-
[25]
S. C. Stearns,The Evolution of Life Histories(Oxford University Press, Oxford, UK, 1992)
work page 1992
- [26]
-
[27]
Virtanenet al., Nature Methods17, 261 (2020)
P. Virtanenet al., Nature Methods17, 261 (2020)
work page 2020
-
[28]
D. T. Gillespie, J. Phys. Chem.81, 2340 (1977). 6 SUPPLEMENT AL MA TERIAL FOR “EMERGENT POPULA TION DYNAMICS OF RANDOM W ALKERS WITH COOPERA TIVE REPRODUCTION AND SP A TIAL SELECTION” Here we provide some details to support the derivations of the main text. In what follows, the notations and abbreviations are the same as in the main text, and the numbered...
work page 1977
-
[29]
Compute the total event rateW tot =W hop +W rxn, whereW hop = 2D 0Nis the total hopping rate of allN particles (D 0 is the per-direction hopping rate) andW rxn = P j wj with the per-site reaction rate (n j is the number of particles at sitej) wj = Λ N k−1 (nj)k ,(n) k ≡n(n−1)· · ·(n−k+1).(S6) Here (n)k is the falling factorial and Λ =λ/k! is the rate cons...
-
[30]
Draw the waiting timeδt=−lnU 1/Wtot, whereU 1 is a uniform random number on (0,1), and advance the clock
-
[31]
With probabilityW hop/Wtot, execute ahop: select a particle uniformly at random and move it to a randomly chosen nearest neighbor
-
[32]
Whenn jL drops to zero,j L advances to the next occupied site
Otherwise, execute areaction-and-removal: select the reaction sitejwith probabilityw j/Wrxn, create a new particle atj, and simultaneously remove one particle from the leftmost occupied sitej L(t). Whenn jL drops to zero,j L advances to the next occupied site. This coupled step conserves the total particle numberNexactly. 8 0 20 40 60 80 z = y/L(t) 0.0 0....
work page 2000
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.