Recognition: unknown
Epistatic strength, modularity, and locus heterogeneity shape the number of local optima in fitness landscapes
Pith reviewed 2026-05-08 01:54 UTC · model grok-4.3
The pith
The expected number of local optima in unstructured fitness landscapes is fixed by the correlation of fitness effects.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
For a broad class of unstructured fitness landscapes modeled as isotropic Gaussian random fields, the expected number of local optima is determined by a single local measure of epistasis: the correlation of fitness effects. This measure links peak density directly to the amount of reciprocal sign epistasis. When epistatic interactions are structured by clustering within blocks of loci, the number of local optima increases slightly; strong heterogeneity that restricts epistasis to a small subset of loci causes the number of peaks to collapse. The results reconcile the central role of reciprocal sign epistasis with the observation that landscapes sharing similar overall epistasis can still be
What carries the argument
The correlation of fitness effects, which quantifies how the fitness impact of a mutation at one locus depends on the state at another and serves as the sole parameter controlling peak density in the Gaussian random-field model.
If this is right
- Stronger negative correlation between fitness effects produces more local optima and therefore more trapping of adaptive walks.
- Clustering epistatic interactions into modular blocks of loci modestly increases the expected number of peaks relative to the unstructured baseline.
- Locus heterogeneity that confines epistasis to a small fraction of sites sharply reduces the number of local optima.
- The model supplies a quantitative baseline for peak numbers in typical unstructured landscapes once the correlation of fitness effects is measured.
- Landscapes with comparable overall epistasis can still differ widely in ruggedness depending on how the interactions are distributed.
Where Pith is reading between the lines
- Measuring correlation of fitness effects in laboratory or natural populations could predict evolutionary accessibility without exhaustive mapping of the full landscape.
- Genomes with high modularity may therefore impose more trapping than random-interaction models suggest.
- Experiments that systematically vary the degree of locus heterogeneity while holding overall epistasis fixed could test whether peak counts collapse as predicted.
Load-bearing premise
Real fitness landscapes can be usefully approximated by isotropic Gaussian random fields in which the correlation of fitness effects fully captures the epistatic structure that matters for counting local optima.
What would settle it
An empirical fitness landscape whose measured correlation of fitness effects predicts a certain number of local optima but whose actual enumeration of peaks deviates substantially from that prediction.
Figures
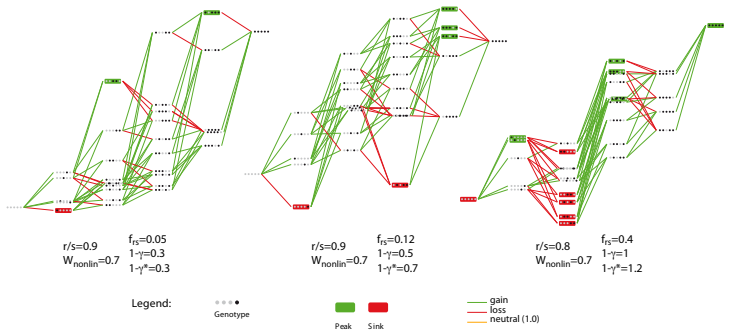

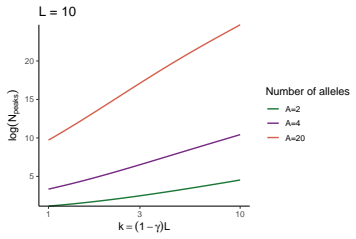



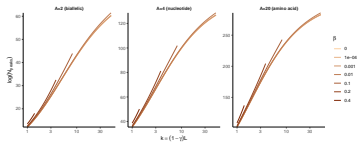
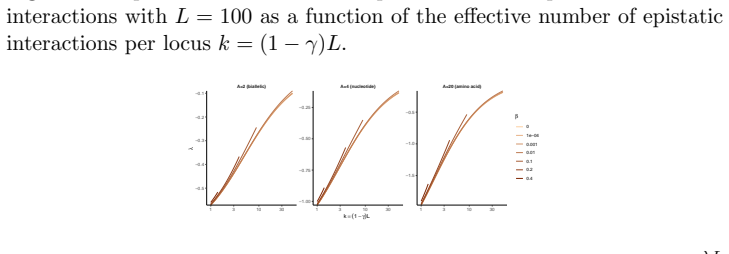
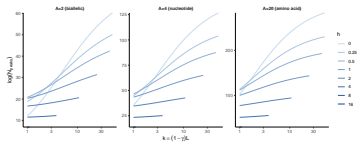

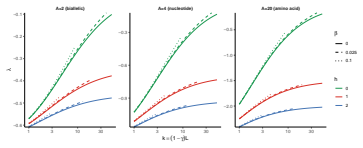
read the original abstract
Fitness landscapes provide a quantitative framework for understanding how natural selection shapes evolutionary trajectories. A central feature of these landscapes is their number of local optima, which determines whether fitness-increasing evolution can proceed towards a global optimum or become trapped on suboptimal peaks. Although multiple peaks are known to require reciprocal sign epistasis, the quantitative relationship between epistasis and number of peaks remains incompletely understood. Here, we show that for a broad class of unstructured fitness landscapes, i.e. isotropic Gaussian random fields, the expected number of local optima is determined by a single local measure of epistasis: the correlation of fitness effects. This provides a baseline prediction for the number of peaks in typical unstructured landscapes and links peak density directly to the amount of reciprocal sign epistasis. This baseline changes when epistatic interactions are structured. We show that clustering interactions within blocks of loci slightly increases the number of local optima. In contrast, strong heterogeneity between loci, where only a small subset of loci participate in epistatic interactions, causes the number of peaks to collapse. These results show that the number of local optima is governed not only by the overall strength of epistasis, but also by how epistatic interactions are distributed across the genotype space. Our framework therefore reconciles the central role of reciprocal sign epistasis with the observation that landscapes with similar amounts of epistasis can differ substantially in ruggedness, and provides a guide to the range of peak numbers expected in typical landscapes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that for isotropic Gaussian random field fitness landscapes on the hypercube, the expected number of local optima equals 2^L times the negative orthant probability of the L-dimensional MVN whose components are the mutational fitness effects, which are exchangeable and equicorrelated with off-diagonal correlation rho determined solely by the covariance at Hamming distances 0, 1 and 2. It further shows that clustering epistatic interactions into modular blocks slightly raises this count while strong locus heterogeneity (only a small subset of loci participating in epistasis) causes the count to collapse, thereby linking peak density to both the overall strength and the spatial distribution of epistasis.
Significance. If the derivations hold, the result supplies a clean, parameter-light baseline for the number of peaks in unstructured landscapes and demonstrates how interaction structure modulates ruggedness even at fixed overall epistasis. The use of linearity of expectation on local-optimality indicators together with the MVN orthant probability is a methodological strength that makes the unstructured case fully determined by a single local statistic.
major comments (2)
- [Main derivation] Main derivation (unstructured case): the claim that only the correlation of fitness effects enters the peak count is correct for isotropic GRFs, but the manuscript should explicitly derive the value of rho from the underlying covariance function evaluated at distances 0, 1 and 2 and state the precise isotropy assumption required for the deltas to remain exchangeable.
- [Structured landscapes] Structured landscapes section: the statements that modularity 'slightly increases' and heterogeneity 'causes collapse' of the peak count need to be accompanied by the exact block sizes, participation fractions, and whether these results follow from an analytic extension of the MVN construction or from Monte Carlo sampling; without these details the quantitative claims cannot be verified.
minor comments (2)
- [Abstract] Abstract: the phrase 'reciprocal sign epistasis' should be tied explicitly to the sign pattern of the vector of mutational effects delta_i(x) in the main text.
- Notation: define the correlation of fitness effects (rho) in a dedicated equation early in the methods so that later references to it are unambiguous.
Simulated Author's Rebuttal
We thank the referee for their positive assessment and constructive suggestions, which will improve the clarity of the derivations and results. We address each major comment below and will incorporate the requested details in the revised manuscript.
read point-by-point responses
-
Referee: Main derivation (unstructured case): the claim that only the correlation of fitness effects enters the peak count is correct for isotropic GRFs, but the manuscript should explicitly derive the value of rho from the underlying covariance function evaluated at distances 0, 1 and 2 and state the precise isotropy assumption required for the deltas to remain exchangeable.
Authors: We agree that an explicit derivation will strengthen the presentation. In the revised manuscript we will add a dedicated paragraph deriving ρ directly from the covariance function evaluated at Hamming distances 0, 1 and 2, and we will state the isotropy assumption (invariance under permutation of loci together with dependence only on Hamming distance) that guarantees exchangeability of the mutational fitness-effect vector. This makes transparent why the orthant probability, and hence the expected number of local optima, depends only on this single local statistic. revision: yes
-
Referee: Structured landscapes section: the statements that modularity 'slightly increases' and heterogeneity 'causes collapse' of the peak count need to be accompanied by the exact block sizes, participation fractions, and whether these results follow from an analytic extension of the MVN construction or from Monte Carlo sampling; without these details the quantitative claims cannot be verified.
Authors: We accept that the current text is insufficiently precise on these points. The modularity and heterogeneity results are obtained by Monte Carlo sampling of the corresponding multivariate normal distributions (not by analytic extension of the unstructured MVN formula). In the revision we will report the exact block sizes (e.g., equal-sized modules of k loci), the participation fractions (e.g., fraction of loci that participate in epistatic interactions), the number of Monte Carlo replicates, and the sampling procedure used to estimate the orthant probabilities. revision: yes
Circularity Check
No significant circularity; derivation is self-contained
full rationale
The paper derives that, for isotropic Gaussian random fields on the hypercube, the expected number of local optima equals 2^L times the negative orthant probability of an L-dimensional MVN whose off-diagonal correlation rho is fixed by the field's covariance at Hamming distances 0, 1 and 2. This follows directly from linearity of expectation applied to the indicator that a genotype is a local optimum (sign pattern of its mutational effects) and the exchangeability of the jointly Gaussian effects; no parameters are fitted to data, no self-citation chain is load-bearing for the central reduction, and the correlation of fitness effects is an input statistic computed from the covariance rather than a quantity defined in terms of the peak count itself. The result is therefore a genuine simplification, not a tautology.
Axiom & Free-Parameter Ledger
free parameters (1)
- correlation of fitness effects
axioms (1)
- domain assumption Fitness landscapes belong to the class of isotropic Gaussian random fields
Reference graph
Works this paper leans on
-
[1]
Journal of Physics A: Mathematical and Theoretical , volume=
Alphabet cardinality and adaptive evolution , author=. Journal of Physics A: Mathematical and Theoretical , volume=. 2023 , publisher=
2023
-
[2]
Biological cybernetics , volume=
Fourier and Taylor series on fitness landscapes , author=. Biological cybernetics , volume=. 1991 , publisher=
1991
-
[3]
SciPost Physics Lecture Notes , pages=
Complexity and accessibility of random landscapes , author=. SciPost Physics Lecture Notes , pages=
-
[4]
, author=
Protein evolution on rugged landscapes. , author=. Proceedings of the National Academy of Sciences , volume=
-
[5]
2012 , publisher=
The adaptive landscape in evolutionary biology , author=. 2012 , publisher=
2012
-
[6]
2026 , eprint=
Evolution as fitness landscape navigation: Concepts, Measures, and Emerging Questions , author=. 2026 , eprint=
2026
-
[7]
PLoS computational biology , volume=
Beyond the hypercube: evolutionary accessibility of fitness landscapes with realistic mutational networks , author=. PLoS computational biology , volume=. 2016 , publisher=
2016
-
[8]
Ferretti, Luca and Schmiegelt, Benjamin and Weinreich, Daniel and Yamauchi, Atsushi and Kobayashi, Yutaka and Tajima, Fumio and Achaz, Guillaume , Date-Added =. Measuring epistasis in fitness landscapes: The correlation of fitness effects of mutations , Volume =. 2016 , Bdsk-Url-1 =. doi:10.1016/j.jtbi.2016.01.037 , Journal =
-
[9]
Evolutionary Accessibility of Modular Fitness Landscapes , Volume =
Schmiegelt, B and Krug, J , Date-Added =. Evolutionary Accessibility of Modular Fitness Landscapes , Volume =. Journal of Statistical Physics , Pages =
-
[10]
A simple stochastic gene substitution model , Volume =
Gillespie, J H , Date-Added =. A simple stochastic gene substitution model , Volume =. Theor Popul Biol , Journal-Full =
-
[11]
1992 , publisher=
Spin Glasses and Biology , editor=. 1992 , publisher=
1992
-
[12]
Spin Glasses and Biology
Stein, Daniel L , Date-Added =. Spin Glasses and Biology. Series: Series on Directions in Condensed Matter Physics, ISBN: 978-9971-5-0537-0. WORLD SCIENTIFIC, Edited by Daniel L Stein, vol. 6 , Title =
-
[13]
Weinreich, Daniel M and Knies, Jennifer L , Date-Added =. Fisher's geometric model of adaptation meets the functional synthesis: data on pairwise epistasis for fitness yields insights into the shape and size of phenotype space , Volume =. 2013 , Bdsk-Url-1 =. doi:10.1111/evo.12156 , Journal =
-
[14]
bioRxiv , Title =
Brouillet, Sophie and Annony, Harry and Ferretti, Luca and Achaz, Guillaume , Date-Added =. bioRxiv , Title =
-
[15]
Should evolutionary geneticists worry about higher-order epistasis? , Volume =
Weinreich, Daniel M and Lan, Yinghong and Wylie, C Scott and Heckendorn, Robert B , Date-Added =. Should evolutionary geneticists worry about higher-order epistasis? , Volume =. 2013 , Bdsk-Url-1 =. doi:10.1016/j.gde.2013.10.007 , Journal =
-
[16]
Annual Review of Ecology, Evolution, and Systematics , Title =
Tenaillon, Olivier , Date-Added =. Annual Review of Ecology, Evolution, and Systematics , Title =
-
[17]
Genetics , volume=
Adaptation in tunably rugged fitness landscapes: the rough Mount Fuji model , author=. Genetics , volume=. 2014 , publisher=
2014
-
[18]
The changing geometry of a fitness landscape along an adaptive walk , Volume =
Greene, Devin and Crona, Kristina , Date-Added =. The changing geometry of a fitness landscape along an adaptive walk , Volume =. 2014 , Bdsk-Url-1 =. doi:10.1371/journal.pcbi.1003520 , Journal =
-
[19]
The Genetical Theory of Natural Selection , Year =
R A Fisher , Date-Added =. The Genetical Theory of Natural Selection , Year =
-
[20]
arXiv , Title =
Blanquart, Fran. arXiv , Title =
-
[21]
Analysis of epistatic interactions and fitness landscapes using a new geometric approach , Volume =
Beerenwinkel, Niko and Pachter, Lior and Sturmfels, Bernd and Elena, Santiago F and Lenski, Richard E , Date-Added =. Analysis of epistatic interactions and fitness landscapes using a new geometric approach , Volume =. 2007 , Bdsk-Url-1 =. doi:10.1186/1471-2148-7-60 , Journal =
-
[22]
The peaks and geometry of fitness landscapes , Volume =
Crona, Kristina and Greene, Devin and Barlow, Miriam , Date-Added =. The peaks and geometry of fitness landscapes , Volume =. 2013 , Bdsk-Url-1 =. doi:10.1016/j.jtbi.2012.09.028 , Journal =
-
[23]
Exact results for amplitude spectra of fitness landscapes , Volume =
Neidhart, Johannes and Szendro, Ivan G and Krug, Joachim , Date-Added =. Exact results for amplitude spectra of fitness landscapes , Volume =. 2013 , Bdsk-Url-1 =. doi:10.1016/j.jtbi.2013.05.002 , Journal =
-
[24]
Selection biases the prevalence and type of epistasis along adaptive trajectories , Volume =
Draghi, Jeremy A and Plotkin, Joshua B , Date-Added =. Selection biases the prevalence and type of epistasis along adaptive trajectories , Volume =. 2013 , Bdsk-Url-1 =. doi:10.1111/evo.12192 , Journal =
-
[25]
Local properties of Kauffman's N-k model: A tunably rugged energy landscape , Volume =
Weinberger, Edward D , Date-Added =. Local properties of Kauffman's N-k model: A tunably rugged energy landscape , Volume =. Physical Review A , Number =
-
[26]
Empirical fitness landscapes reveal accessible evolutionary paths , Volume =
Poelwijk, Frank J and Kiviet, Daniel J and Weinreich, Daniel M and Tans, Sander J , Date-Added =. Empirical fitness landscapes reveal accessible evolutionary paths , Volume =. 2007 , Bdsk-Url-1 =. doi:10.1038/nature05451 , Journal =
-
[27]
A cross-section of the fitness landscape of dihydrofolate reductase , Volume =
Aita, T and Iwakura, M and Husimi, Y , Date-Added =. A cross-section of the fitness landscape of dihydrofolate reductase , Volume =. Protein Eng , Journal-Full =
-
[28]
Landscapes and their correlation functions , Volume =
Stadler, Peter F , Date-Added =. Landscapes and their correlation functions , Volume =. Journal of Mathematical chemistry , Number =
-
[29]
Analysis of a local fitness landscape with a model of the rough Mt
Aita, T and Uchiyama, H and Inaoka, T and Nakajima, M and Kokubo, T and Husimi, Y , Date-Added =. Analysis of a local fitness landscape with a model of the rough Mt. Fuji-type landscape: application to prolyl endopeptidase and thermolysin , Volume =. 2000 , Bdsk-Url-1 =. doi:10.1002/(SICI)1097-0282(200007)54:1<64::AID-BIP70>3.0.CO;2-R , Journal =
-
[30]
Achaz, Guillaume and Rodriguez-Verdugo, Alejandra and Gaut, Brandon S and Tenaillon, Olivier , Date-Added =. The reproducibility of adaptation in the light of experimental evolution with whole genome sequencing , Volume =. 2014 , Bdsk-Url-1 =. doi:10.1007/978-94-007-7347-9_11 , Journal =
-
[31]
The NK model of rugged fitness landscapes and its application to maturation of the immune response , Volume =
Kauffman, S A and Weinberger, E D , Date-Added =. The NK model of rugged fitness landscapes and its application to maturation of the immune response , Volume =. J Theor Biol , Journal-Full =
-
[32]
Spin glass theory and beyond , Year =
M. Spin glass theory and beyond , Year =
-
[33]
Visualizing fitness landscapes , Volume =
McCandlish, David M , Date-Added =. Visualizing fitness landscapes , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1111/j.1558-5646.2011.01236.x , Journal =
-
[34]
The genetical theory of natural selection , Year =
Fisher, RA , Date-Added =. The genetical theory of natural selection , Year =
-
[35]
On the existence of accessible paths in various models of fitness landscapes , Volume =
Hegarty, Peter and Martinsson, Anders and others , Date-Added =. On the existence of accessible paths in various models of fitness landscapes , Volume =. The Annals of Applied Probability , Number =
-
[36]
arXiv preprint arXiv:1304.0246 , Title =
Berestycki, Julien and Brunet,. arXiv preprint arXiv:1304.0246 , Title =
-
[37]
Towards a general theory of adaptive walks on rugged landscapes , Volume =
Kauffman, S and Levin, S , Date-Added =. Towards a general theory of adaptive walks on rugged landscapes , Volume =. J Theor Biol , Journal-Full =
-
[38]
A simple model for the balance between selection and mutation , Year =
Kingman, JFC , Date-Added =. A simple model for the balance between selection and mutation , Year =. Journal of Applied Probability , Pages =
-
[39]
On the genealogy of large populations , Year =
Kingman, John FC , Date-Added =. On the genealogy of large populations , Year =. Journal of Applied Probability , Pages =
-
[40]
Reciprocal sign epistasis is a necessary condition for multi-peaked fitness landscapes , Volume =
Poelwijk, Frank J and T. Reciprocal sign epistasis is a necessary condition for multi-peaked fitness landscapes , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1016/j.jtbi.2010.12.015 , Journal =
-
[41]
Initial mutations direct alternative pathways of protein evolution , Volume =
Salverda, Merijn L M and Dellus, Eynat and Gorter, Florien A and Debets, Alfons J M and van der Oost, John and Hoekstra, Rolf F and Tawfik, Dan S and de Visser, J Arjan G M , Date-Added =. Initial mutations direct alternative pathways of protein evolution , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1371/journal.pgen.1001321 , Journal =
-
[42]
Chevin, Luis-Miguel and Martin, Guillaume and Lenormand, Thomas , Date-Added =. Fisher's model and the genomics of adaptation: restricted pleiotropy, heterogenous mutation, and parallel evolution , Volume =. 2010 , Bdsk-Url-1 =. doi:10.1111/j.1558-5646.2010.01058.x , Journal =
-
[43]
Microbial experiments on adaptive landscapes , Volume =
Colegrave, Nick and Buckling, Angus , Date-Added =. Microbial experiments on adaptive landscapes , Volume =. 2005 , Bdsk-Url-1 =. doi:10.1002/bies.20292 , Journal =
-
[44]
The evolutionary enigma of sex , Volume =
Otto, Sarah P , Date-Added =. The evolutionary enigma of sex , Volume =. 2009 , Bdsk-Url-1 =. doi:10.1086/599084 , Journal =
-
[45]
Exploring the effect of sex on empirical fitness landscapes , Volume =
de Visser, J Arjan G M and Park, Su-Chan and Krug, Joachim , Date-Added =. Exploring the effect of sex on empirical fitness landscapes , Volume =. 2009 , Bdsk-Url-1 =. doi:10.1086/599081 , Journal =
-
[46]
Optimal adaptive performance and delocalization in NK fitness landscapes , Volume =
Campos, Paulo RA and Adami, Christoph and Wilke, Claus O , Date-Added =. Optimal adaptive performance and delocalization in NK fitness landscapes , Volume =. Physica A: Statistical Mechanics and its Applications , Number =
-
[47]
Fitness Landscapes: From Evolutionary Biology to Evolutionary Computation , Year =
Richter, Hendrik , Booktitle =. Fitness Landscapes: From Evolutionary Biology to Evolutionary Computation , Year =
-
[48]
Random-energy model: Limit of a family of disordered models , Volume =
Derrida, Bernard , Date-Added =. Random-energy model: Limit of a family of disordered models , Volume =. Physical Review Letters , Number =
-
[49]
The genetic theory of adaptation: a brief history , Volume =
Orr, H Allen , Date-Added =. The genetic theory of adaptation: a brief history , Volume =. 2005 , Bdsk-Url-1 =. doi:10.1038/nrg1523 , Journal =
-
[50]
Empirical fitness landscapes and the predictability of evolution , Volume =
de Visser, J Arjan G M and Krug, Joachim , Date-Added =. Empirical fitness landscapes and the predictability of evolution , Volume =. 2014 , Bdsk-Url-1 =. doi:10.1038/nrg3744 , Journal =
-
[51]
Watson, Richard A and Weinreich, Daniel M and Wakeley, John , Date-Added =. Genome structure and the benefit of sex , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1111/j.1558-5646.2010.01144.x , Journal =
-
[52]
Niche dimensionality and the genetics of ecological speciation , Volume =
Chevin, Luis-Miguel and Decorzent, Guillaume and Lenormand, Thomas , Date-Added =. Niche dimensionality and the genetics of ecological speciation , Volume =. 2014 , Bdsk-Url-1 =. doi:10.1111/evo.12346 , Journal =
-
[53]
Colloquium papers: Adaptive landscapes and protein evolution , Volume =
Carneiro, Maur. Colloquium papers: Adaptive landscapes and protein evolution , Volume =. 2010 , Bdsk-Url-1 =. doi:10.1073/pnas.0906192106 , Journal =
-
[54]
Test of interaction between genetic markers that affect fitness in Aspergillus niger
de Visser, J A and Hoekstra, R F and van den Ende, H , Date-Added =. Test of interaction between genetic markers that affect fitness in Aspergillus niger. , Volume =. Evolution , Pages =
-
[55]
Evolutionary accessibility of mutational pathways , Volume =
Franke, Jasper and Kl. Evolutionary accessibility of mutational pathways , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1371/journal.pcbi.1002134 , Journal =
-
[56]
Fitness Landscapes and the Origin of Species , Year =
Sergey Gavrilets , Date-Added =. Fitness Landscapes and the Origin of Species , Year =
-
[57]
The Causes of Molecular Evolution , Year =
Gillespie, J H , Date-Added =. The Causes of Molecular Evolution , Year =
-
[58]
Bio Philos , Title =
Kaplan, J , Date-Added =. Bio Philos , Title =
-
[59]
The Origins of Order: Self-Organization and Selection in Evolution , Year =
Stuart Kauffman , Date-Added =. The Origins of Order: Self-Organization and Selection in Evolution , Year =
-
[60]
Negative epistasis between beneficial mutations in an evolving bacterial population , Volume =
Khan, Aisha I and Dinh, Duy M and Schneider, Dominique and Lenski, Richard E and Cooper, Tim F , Date-Added =. Negative epistasis between beneficial mutations in an evolving bacterial population , Volume =. 2011 , Bdsk-Url-1 =. doi:10.1126/science.1203801 , Journal =
-
[61]
The Neutral Theory of Molecular Evolution , Year =
Kimura, Motoo , Date-Added =. The Neutral Theory of Molecular Evolution , Year =
-
[62]
Multidimensional epistasis and the disadvantage of sex , Volume =
Kondrashov, F A and Kondrashov, A S , Date-Added =. Multidimensional epistasis and the disadvantage of sex , Volume =. 2001 , Bdsk-Url-1 =. doi:10.1073/pnas.211214298 , Journal =
-
[63]
Dobzhansky-Muller incompatibilities in protein evolution , Volume =
Kondrashov, Alexey S and Sunyaev, Shamil and Kondrashov, Fyodor A , Date-Added =. Dobzhansky-Muller incompatibilities in protein evolution , Volume =. 2002 , Bdsk-Url-1 =. doi:10.1073/pnas.232565499 , Journal =
-
[64]
Phillips, Patrick C , Date-Added =. Epistasis--the essential role of gene interactions in the structure and evolution of genetic systems , Volume =. 2008 , Bdsk-Url-1 =. doi:10.1038/nrg2452 , Journal =
-
[65]
Journal of Statistical Mechanics: Theory and Experiment , Title =
Szendro, Ivan G and Schenk, Martijn F and Franke, J and Krug, Joachim and de Visser, J Arjan G M , Date-Added =. Journal of Statistical Mechanics: Theory and Experiment , Title =
-
[66]
The molecular diversity of adaptive convergence , Volume =
Tenaillon, Olivier and Rodr. The molecular diversity of adaptive convergence , Volume =. 2012 , Bdsk-Url-1 =. doi:10.1126/science.1212986 , Journal =
-
[67]
Perspective: Sign epistasis and genetic constraint on evolutionary trajectories , Volume =
Weinreich, Daniel M and Watson, Richard A and Chao, Lin , Date-Added =. Perspective: Sign epistasis and genetic constraint on evolutionary trajectories , Volume =. Evolution , Journal-Full =
-
[68]
Weinreich, Daniel M , Date-Added =. The rank ordering of genotypic fitness values predicts genetic constraint on natural selection on landscapes lacking sign epistasis , Volume =. 2005 , Bdsk-Url-1 =. doi:10.1534/genetics.104.036830 , Journal =
-
[69]
Darwinian evolution can follow only very few mutational paths to fitter proteins , Volume =
Weinreich, Daniel M and Delaney, Nigel F and Depristo, Mark A and Hartl, Daniel L , Date-Added =. Darwinian evolution can follow only very few mutational paths to fitter proteins , Volume =. 2006 , Bdsk-Url-1 =. doi:10.1126/science.1123539 , Journal =
-
[70]
Different trajectories of parallel evolution during viral adaptation , Volume =
Wichman, H A and Badgett, M R and Scott, L A and Boulianne, C M and Bull, J J , Date-Added =. Different trajectories of parallel evolution during viral adaptation , Volume =. Science , Journal-Full =
-
[71]
The roles of mutation, inbreeding, crossbreeding and selection in evolution , Volume =
S Wright , Date-Added =. The roles of mutation, inbreeding, crossbreeding and selection in evolution , Volume =. Proc. 6th Int. Cong. Genet. , Pages =
-
[72]
Nature , volume=
Ancestral lysozymes reconstructed, neutrality tested and thermostability linked to hydrocarbon packing , author=. Nature , volume=. 1990 , publisher=
1990
-
[73]
Evolution , volume=
Factors affecting the genetic load in Drosophila: synergistic epistasis and correlations among fitness components , author=. Evolution , volume=. 2000 , publisher=
2000
-
[74]
Science , volume=
The biochemical architecture of an ancient adaptive landscape , author=. Science , volume=. 2005 , publisher=
2005
-
[75]
Nature chemical biology , volume=
Quantitative exploration of the catalytic landscape separating divergent plant sesquiterpene synthases , author=. Nature chemical biology , volume=. 2008 , publisher=
2008
-
[76]
Nature , volume=
An epistatic ratchet constrains the direction of glucocorticoid receptor evolution , author=. Nature , volume=. 2009 , publisher=
2009
-
[77]
Proceedings of the National Academy of Sciences , volume=
Stepwise acquisition of pyrimethamine resistance in the malaria parasite , author=. Proceedings of the National Academy of Sciences , volume=. 2009 , publisher=
2009
-
[78]
Journal of heredity , volume=
Fitness epistasis among 6 biosynthetic loci in the budding yeast Saccharomyces cerevisiae , author=. Journal of heredity , volume=. 2010 , publisher=
2010
-
[79]
Genetics , volume=
Fitness epistasis and constraints on adaptation in a human immunodeficiency virus type 1 protein region , author=. Genetics , volume=. 2010 , publisher=
2010
-
[80]
Physical review letters , volume=
Hidden randomness between fitness landscapes limits reverse evolution , author=. Physical review letters , volume=. 2011 , publisher=
2011
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.