Recognition: unknown
Chapter 2: Geometry of the Fitness Surface and Trajectory Dynamics of Replicator Systems
Pith reviewed 2026-05-08 15:45 UTC · model grok-4.3
The pith
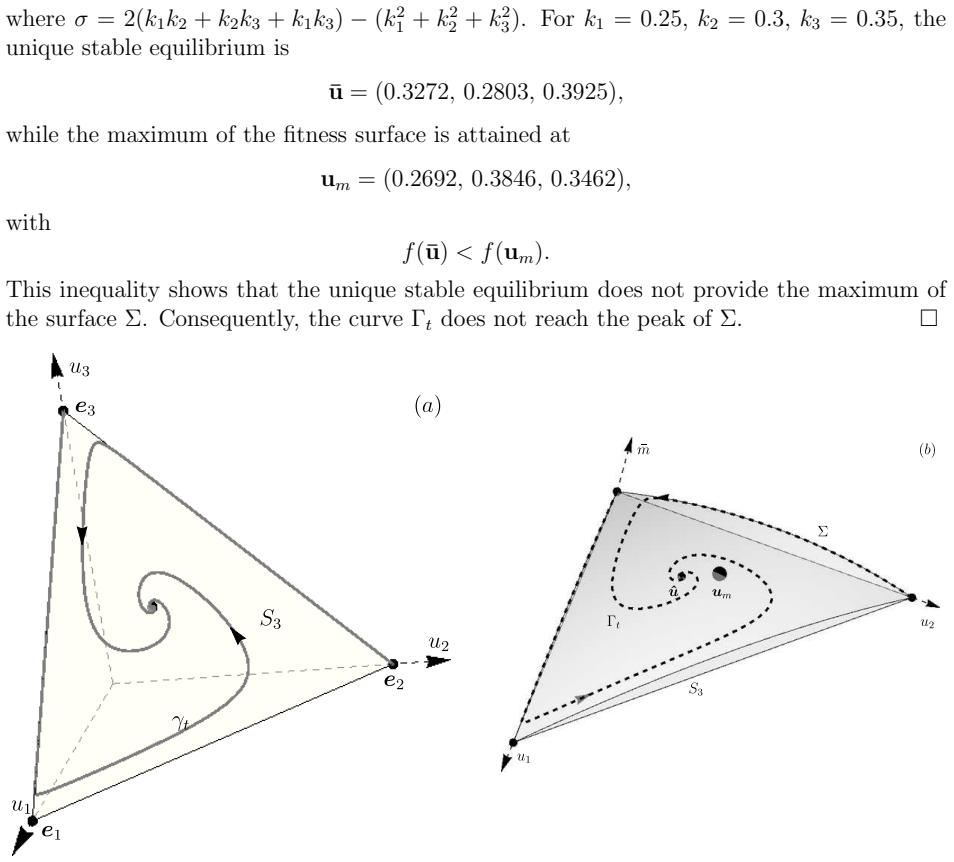
Replicator trajectories do not reach the maximum of the fitness surface unless the equilibrium is a local extremum characterized by the symmetric and antisymmetric parts of the fitness matrix.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Using the symmetric-antisymmetric decomposition of the fitness landscape matrix, we derive an explicit formula for the rate of change of mean fitness and establish necessary conditions for its monotonicity along trajectories. In general, replicator trajectories do not reach the maximum of the fitness surface, even in the presence of a unique asymptotically stable equilibrium. We characterise the precise conditions under which an equilibrium coincides with a local extremum of the fitness surface in terms of the symmetric and antisymmetric parts of the fitness matrix, with circulant matrices serving as a natural example. Evolutionary stability implies a local fitness maximum, and the converse
What carries the argument
The symmetric-antisymmetric decomposition of the fitness matrix, which separates the quadratic form that governs changes in mean fitness from the skew-symmetric component that shapes trajectory paths on the simplex.
Load-bearing premise
The fitness landscape is represented by a fixed matrix that admits a symmetric-antisymmetric decomposition and the dynamics follow the standard replicator equation on the probability simplex without additional nonlinearities, time dependence, or stochastic effects.
What would settle it
A replicator system with a unique asymptotically stable equilibrium that is not a local extremum of the mean fitness surface, or a trajectory that reaches the global maximum without the equilibrium satisfying the characterized conditions on the matrix decomposition.
Figures

read the original abstract
We study the geometry of the mean fitness surface of replicator systems and its relationship to evolutionary trajectory dynamics. Using the symmetric--antisymmetric decomposition of the fitness landscape matrix, we derive an explicit formula for the rate of change of mean fitness and establish necessary conditions for its monotonicity along trajectories. In general, replicator trajectories do not reach the maximum of the fitness surface, even in the presence of a unique asymptotically stable equilibrium. We characterise, in terms of the symmetric and antisymmetric parts of the fitness matrix, the precise conditions under which an equilibrium coincides with a local extremum of the fitness surface. Circulant matrices are identified as a natural and nontrivial class satisfying these conditions. We establish a two-way connection between fitness surface maxima and evolutionarily stable states: evolutionary stability implies a local fitness maximum, and the converse holds under the identified structural conditions. When the unique asymptotically stable equilibrium is a local maximum, it is evolutionarily stable and realises the global maximum of the fitness surface; an unstable equilibrium forces the global maximum to the boundary of the simplex. The framework is extended to general Lotka--Volterra systems, where an analogue of mean fitness is shown to share the same extremal properties. Results are illustrated through six examples spanning autocatalytic and hypercyclic replication, a parametric family exhibiting Andronov--Hopf bifurcation and heteroclinic cycles, and the Eigen quasispecies model.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript analyzes the geometry of the mean fitness surface for replicator systems via the symmetric-antisymmetric decomposition A = S + K of the fitness matrix. It derives an explicit formula for d(mean fitness)/dt along trajectories of the replicator equation, establishes necessary conditions for monotonicity, shows that trajectories generally fail to reach the global fitness maximum even with a unique asymptotically stable equilibrium, and characterizes the precise conditions (in terms of S and K) under which an equilibrium coincides with a local extremum of the fitness surface. Circulant matrices are identified as a natural class satisfying these conditions. A two-way link is established between local fitness maxima and evolutionary stability, with further results on global maxima and boundary behavior. The framework is extended to general Lotka-Volterra systems, where an analogue of mean fitness shares the same extremal properties. Results are illustrated with six examples covering autocatalytic and hypercyclic replication, a parametric family with Andronov-Hopf bifurcation and heteroclinic cycles, and the Eigen quasispecies model.
Significance. If the derivations hold, the work supplies a clean geometric and variational perspective on fitness landscapes in evolutionary game theory. The decomposition cleanly separates the non-negative variance contribution from S and the uncontrolled skew term from K, yielding explicit monotonicity conditions and a precise characterization of when equilibria align with extrema. The circulant-matrix class, the bidirectional ESS-maximum link, and the Lotka-Volterra extension are concrete, usable contributions. The six examples provide concrete verification across qualitatively different regimes. These results clarify why mean fitness need not increase to a global maximum and when stability implies extremality, strengthening the analytical toolkit for replicator and related dynamical systems.
minor comments (2)
- The abstract packs many distinct claims into a single paragraph; separating the main results (formula, monotonicity conditions, extremum characterization, ESS link, LV extension) into a short enumerated list would improve immediate readability.
- In the examples section, ensure that all six models are explicitly labeled (e.g., Example 1, Example 2, …) with their specific matrix entries or parameter ranges stated in a self-contained manner so that the illustrated behaviors (monotonicity, Hopf cycles, etc.) can be reproduced without external references.
Simulated Author's Rebuttal
We thank the referee for their positive and accurate summary of the manuscript, for highlighting its contributions to the geometric analysis of fitness surfaces in replicator dynamics, and for recommending minor revision. We appreciate the recognition of the symmetric-antisymmetric decomposition, the monotonicity conditions, the circulant-matrix class, the ESS-maximum link, and the Lotka-Volterra extension. No specific major comments or requests for clarification were raised in the report.
Circularity Check
No significant circularity detected
full rationale
The paper's central derivations compute the time derivative of mean fitness xᵀAx along replicator trajectories using the decomposition A = S + K into symmetric and antisymmetric parts. This yields an explicit expression separating a non-negative variance term (from S) and an uncontrolled skew term (from K), from which necessary conditions for monotonicity follow by direct differentiation and standard variational arguments on the simplex. Equivalence between equilibria and local extrema of the fitness surface, the special role of circulant matrices, and the two-way link to evolutionary stability are all algebraic consequences of these identities and Lyapunov analysis; the Lotka–Volterra extension uses an analogous quadratic form. No fitted parameters are renamed as predictions, no self-definitional loops appear, and no load-bearing self-citations or imported uniqueness theorems are invoked. The claimed results are therefore independent mathematical consequences of the stated assumptions rather than reductions to the inputs by construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The fitness landscape is given by a constant real matrix that admits the decomposition A = S + K with S symmetric and K antisymmetric.
Reference graph
Works this paper leans on
-
[1]
Bratus, A. S., Drozhzhin, S., & Yakushkina, T. Mathematical mod els of evolution and replicator systems dynamics. Chapter 1: Introduction to rep licator systems. arXiv:2604.05720, 2026. https://arxiv.org/abs/2604.05720
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[2]
Fisher, R. A. The Genetical Theory of Natural Selection . Clarendon Press, Oxford, 1930
1930
-
[3]
Crow, J. F. Here’s to Fisher, additive genetic variance, and the f unda- mental theorem of natural selection. Evolution, 56(7):1313–1316, 2002. doi:10.1111/j.0014-3820.2002.tb01443.x
-
[4]
On the change of population fitness by natural select ion
Kimura, M. On the change of population fitness by natural select ion. Heredity, 12(2):145– 167, 1958. doi:10.1038/hdy.1958.15
-
[5]
P., & Lefschetz, S
LaSalle, J. P., & Lefschetz, S. Stability by Liapunov’s Direct Method with Applications . Academic Press, New York, 1961
1961
-
[6]
S., Novozhilov, A
Bratus, A. S., Novozhilov, A. S., & Platonov, A. P. Dynamical Systems and Models in Biology. Fizmatlit, Moscow, 2010. [in Russian]
2010
-
[7]
Evolution and the Theory of Games
Maynard Smith, J. Evolution and the Theory of Games . Cambridge University Press, Cambridge, 1982
1982
-
[8]
The roles of mutation, inbreeding, crossbreeding and selection in evolution
Wright, S. The roles of mutation, inbreeding, crossbreeding and selection in evolution. Proceedings of the Sixth International Congress of Genetic s, 1:356–366, 1932
1932
-
[9]
Evolutionary game dynamics.Bulletin of the American Mathematical Society, 40(4):479–519, 2003
Hofbauer, J., & Sigmund, K. Evolutionary game dynamics. Bulletin of the American Mathematical Society, 40(4):479–519, 2003. doi:10.1090/S0273-0979-03-00988-1 20
-
[10]
D., & Tikhomirov, V
Ioffe, A. D., & Tikhomirov, V. M. Theory of Extremal Problems . Nauka, Moscow, 1974. [in Russian]
1974
-
[11]
A note on evolutionar ily stable strate- gies and game dynamics
Hofbauer, J., Schuster, P., & Sigmund, K. A note on evolutionar ily stable strate- gies and game dynamics. Journal of Theoretical Biology , 81(3):609–612, 1979. doi:10.1016/0022-5193(79)90058-4
-
[12]
Maynard Smith, J., & Price, G. R. The logic of animal conflict. Nature, 246:15–18, 1973. doi:10.1038/246015a0
-
[13]
Drozhzhin, S., Yakushkina, T., & Bratus, A. S. Fitness optimizat ion and evolution of permanent replicator systems. Journal of Mathematical Biology , 82(3):15, 2021. doi:10.1007/s00285-021-01548-8
-
[14]
Naturwissenschaften 1971, 58(10), 465–523
Eigen, M. Selforganization of matter and the evolution of biologic al macromolecules. Naturwissenschaften, 58(10):465–523, 1971. doi:10.1007/BF00623322
-
[15]
M., & Logofet, D
Svirezhev, Yu. M., & Logofet, D. O. Ustoychivost’ biologicheskikh soobshchestv [Stability of Biological Communities]. Nauka, Moscow, 1978. [in Russian] 21
1978
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.