Recognition: unknown
Planar morphometry via functional shape data analysis and quasi-conformal mappings
Pith reviewed 2026-05-08 03:14 UTC · model grok-4.3
The pith
Combining functional boundary registration with quasi-conformal interior extension captures morphological variation in planar shapes more effectively than boundary-only or interior-only methods.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
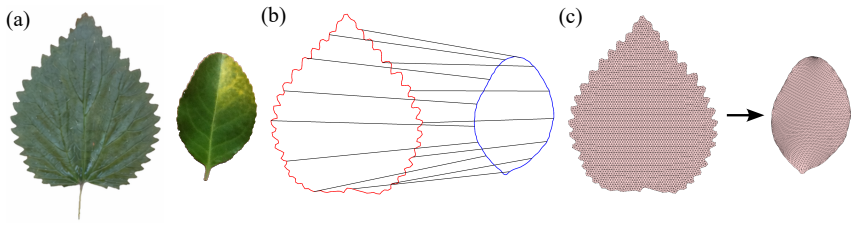
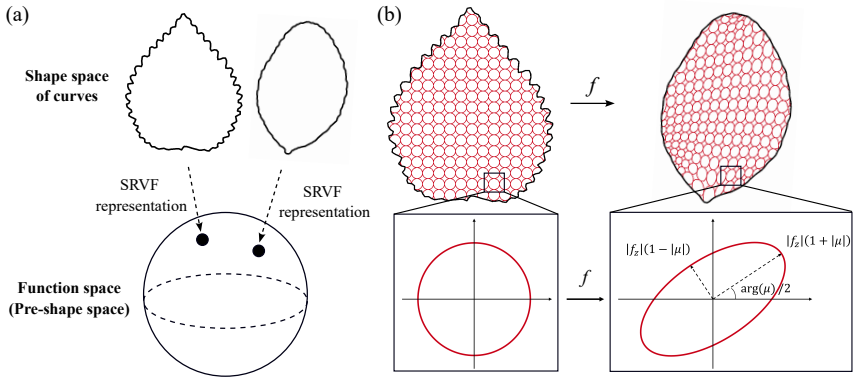
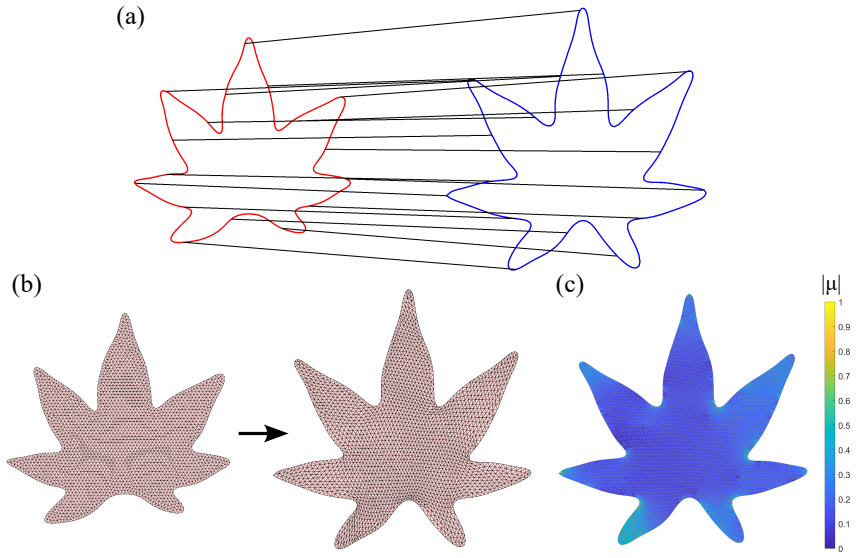
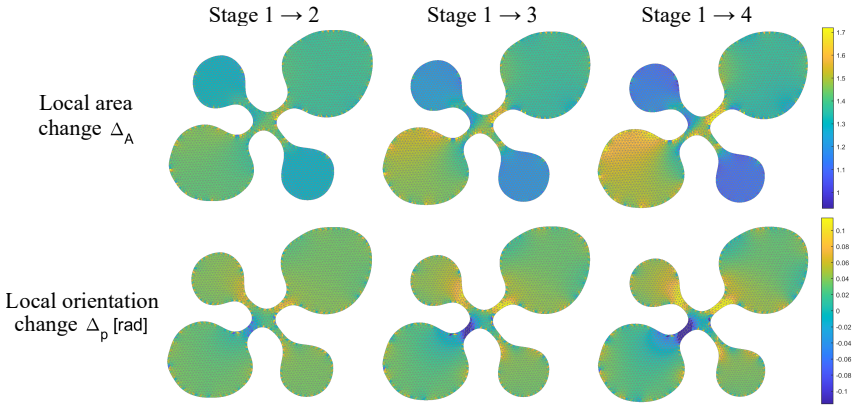
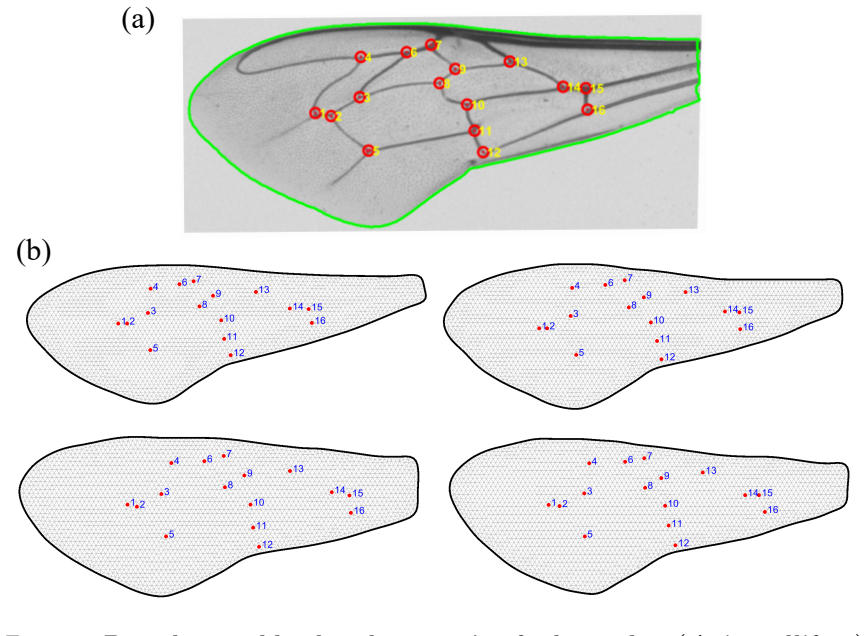
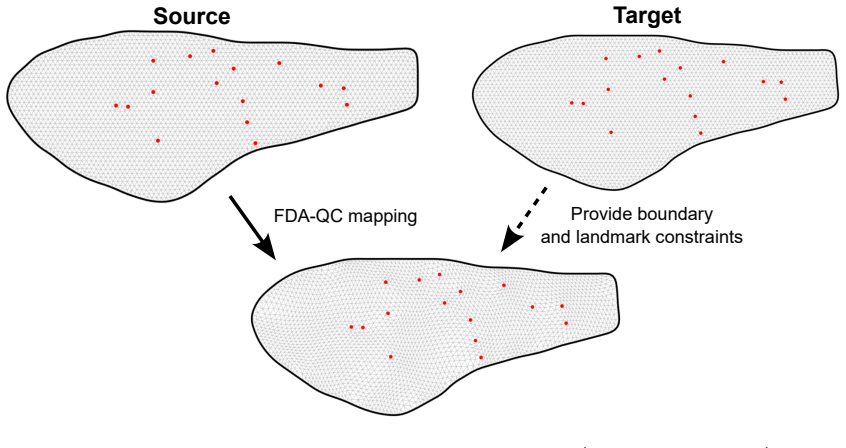
Closed planar curves are represented by their square-root velocity functions and registered via elastic matching; the resulting boundary correspondence is extended to the interior domain by a quasi-conformal map. The combined representation yields a single framework for shape morphing and for quantifying shape variation, and this framework is shown to outperform purely boundary-based or purely interior-based descriptions when applied to collections of leaves and insect wings.
What carries the argument
Quasi-conformal extension of boundary correspondences obtained from square-root velocity function elastic matching
If this is right
- The method supplies a single pipeline that simultaneously registers shapes, morphs them, and quantifies their variation.
- Morphological differences in biological planar forms can be localized to both outline and interior regions in the same coordinate system.
- Landmark constraints can be added or removed without rebuilding the entire registration step.
- Shape variation statistics become comparable across different collections once mapped into the same quasi-conformal coordinate frame.
Where Pith is reading between the lines
- The same boundary-to-interior extension step could be reused as a preprocessing stage for machine-learning classifiers that operate on planar biological images.
- If the quasi-conformal maps remain bijective under moderate noise, the framework could support longitudinal studies that track growth trajectories of individual leaves or wings.
- The elastic matching step in function space might be replaced by other curve-registration techniques while preserving the downstream quasi-conformal extension.
Load-bearing premise
That extending boundary correspondences via quasi-conformal mappings faithfully captures the coupling between boundary and interior features without introducing geometric distortions that affect downstream variation analysis.
What would settle it
A controlled dataset of planar shapes with known interior-boundary feature couplings where the FDA-QC variation measures fail to separate groups better than boundary-only or interior-only measures according to a standard statistical test such as Hotelling's T-squared.
Figures



read the original abstract
The study of shapes is one of the most fundamental problems in life sciences. Although numerous methods have been developed for the morphometry of planar biological shapes over the past several decades, most of them focus solely on either the outer silhouettes or the interior features of the shapes without capturing the coupling between them. Moreover, many existing shape mapping techniques are limited to establishing correspondence between planar structures without further allowing for the quantitative analysis or modelling of shape changes. In this work, we introduce FDA-QC, a novel planar morphometry method that combines functional shape data analysis (FDA) techniques and quasi-conformal (QC) mappings, taking both the boundary and interior of the planar shapes into consideration. Specifically, closed planar curves are represented by their square-root velocity functions and registered by elastic matching in the function space. The induced boundary correspondence is then extended to the entire planar domains by a quasi-conformal map, optionally with landmark constraints. Moreover, the proposed FDA-QC method can naturally lead to a unified framework for shape morphing and shape variation quantification. We apply the FDA-QC method to various leaf and insect wing datasets, and the experimental results show that the proposed combined approach captures morphological variation more effectively than purely boundary-based or interior-based descriptions. Altogether, our work paves a new way for understanding the growth and form of planar biological shapes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces FDA-QC, a planar morphometry framework that registers closed curves via square-root velocity functions (SRVF) and elastic matching in function space, then extends the resulting boundary correspondences to the full planar domain using quasi-conformal mappings (optionally with landmark constraints). The combined representation is used for shape morphing and for quantifying morphological variation; experiments on leaf and insect-wing datasets are reported to show that this boundary-interior coupling captures variation more effectively than purely boundary-based (SRVF) or interior-based descriptions.
Significance. If the superiority claim is supported by quantitative validation, the work would be significant for biological shape analysis: it supplies a unified, extensible pipeline that links established FDA tools with QC theory to model both silhouette and interior features simultaneously, addressing a recognized gap in planar morphometry. The approach also yields a natural morphing mechanism, which is a practical strength.
major comments (2)
- [§3] §3 (quasi-conformal extension): no bound or sensitivity analysis is given for the Beltrami coefficient of the QC map, nor is it shown how any local stretching/compression propagates into the downstream variation measures (e.g., functional PCA or distance on the extended fields). Without this, the central claim that the combined representation “captures morphological variation more effectively” cannot be separated from possible mapping-induced artifacts.
- [§4] §4 (experiments): the superiority of FDA-QC over SRVF-only and interior-only baselines is asserted on leaf and wing data, yet the text supplies no numerical metrics (explained variance, reconstruction error, classification accuracy, or statistical tests) and appears to rely on visual inspection of morphs and variation plots. This renders the effectiveness claim unsupported at the level required for the central result.
minor comments (2)
- Notation for the extended map and the variation functional should be introduced with a single consistent symbol set; current usage mixes curve and domain variables without explicit cross-reference.
- Figure captions for the morphing sequences should state the exact number of landmarks (if any) and the Beltrami-coefficient range used in each example.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback, which highlights important areas for strengthening the manuscript. We address each major comment below and have revised the manuscript to incorporate quantitative analyses and additional discussion as suggested.
read point-by-point responses
-
Referee: §3 (quasi-conformal extension): no bound or sensitivity analysis is given for the Beltrami coefficient of the QC map, nor is it shown how any local stretching/compression propagates into the downstream variation measures (e.g., functional PCA or distance on the extended fields). Without this, the central claim that the combined representation “captures morphological variation more effectively” cannot be separated from possible mapping-induced artifacts.
Authors: We agree that an explicit analysis of the Beltrami coefficient and its influence on downstream measures would strengthen the separation of morphological signal from mapping effects. In the revised manuscript, we have added a new subsection in §3 that reports the distribution of |μ| values across the datasets (all |μ| < 0.35, with mean |μ| ≈ 0.12), confirming the maps remain close to conformal. We also include a sensitivity study: small random perturbations to the boundary correspondence (within the elastic matching tolerance) are propagated through the QC extension, and the resulting changes in functional PCA scores and pairwise distances are quantified and shown to be negligible relative to inter-shape variation. These additions directly address the concern while preserving the original framework. revision: yes
-
Referee: §4 (experiments): the superiority of FDA-QC over SRVF-only and interior-only baselines is asserted on leaf and wing data, yet the text supplies no numerical metrics (explained variance, reconstruction error, classification accuracy, or statistical tests) and appears to rely on visual inspection of morphs and variation plots. This renders the effectiveness claim unsupported at the level required for the central result.
Authors: We acknowledge that the original experimental section relied primarily on qualitative visualization. In the revised manuscript, we have added quantitative comparisons in §4: (i) cumulative explained variance by the first 5 principal components for FDA-QC versus SRVF and interior-only representations; (ii) average reconstruction error (L2 norm on the extended fields) for morphing between pairs of shapes; (iii) leave-one-out classification accuracy for species discrimination using the shape representations as features; and (iv) paired statistical tests (Wilcoxon signed-rank) confirming significant improvement (p < 0.01) of FDA-QC over baselines on both datasets. These metrics are now reported in new tables and support the effectiveness claim with objective evidence. revision: yes
Circularity Check
No significant circularity; method combines established FDA and QC techniques with independent empirical validation
full rationale
The paper's core contribution is a hybrid FDA-QC pipeline: SRVF elastic registration of boundaries (standard in functional data analysis literature) followed by quasi-conformal extension to the interior (standard in QC mapping literature), optionally with landmarks. The claimed superiority in capturing morphological variation is demonstrated via direct experimental comparison on leaf and insect wing datasets against purely boundary-based (SRVF) and interior-based baselines. No equation or result reduces by construction to a fitted parameter, self-definition, or self-citation chain; the variation quantification follows from the composed maps without tautological equivalence to the input correspondences. Self-citations to prior FDA/QC works are not load-bearing for the central empirical claim, which remains falsifiable against the reported baselines.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Quasi-conformal mappings can extend a boundary correspondence to the interior domain while respecting optional landmark constraints.
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.