Recognition: no theorem link
Accounting for Missed Events in the Bayesian Modeling of IP3R Multimodal Gating
Pith reviewed 2026-05-13 00:52 UTC · model grok-4.3
The pith
Integrating missed-event correction into Bayesian likelihood clarifies IP3R's Park and Drive gating modes as variants of a single 3-state Markov model.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
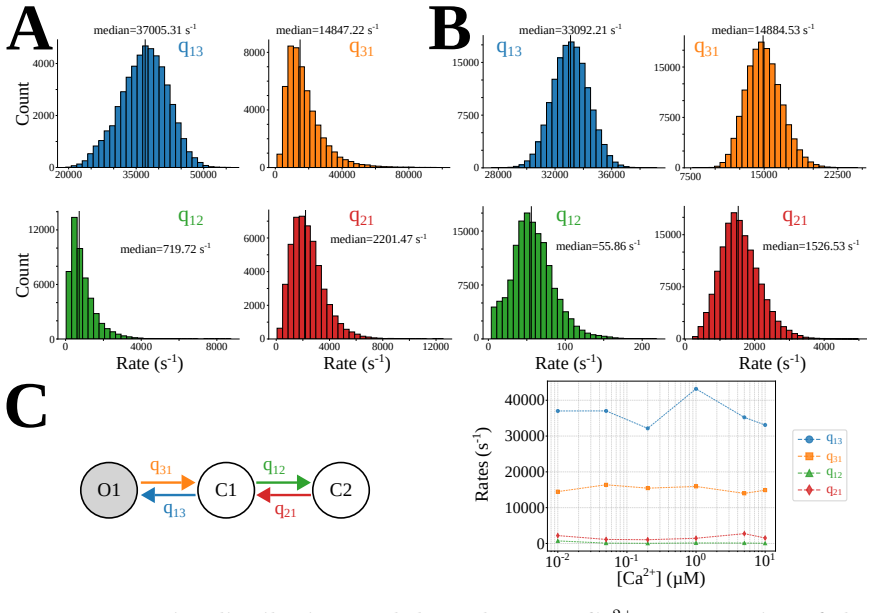
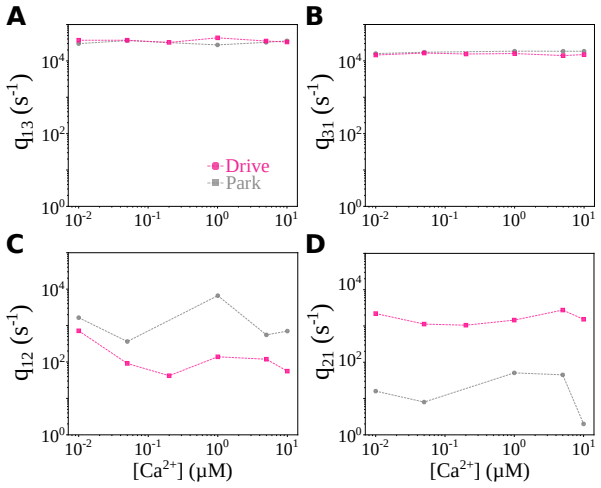
By incorporating a correction for missed events directly into the likelihood function within a Bayesian framework for hierarchical Markov chain models, the selected model shows that IP3R gating operates in Park and Drive modes that each correspond to the same 3-state Markov model, but with mode-dependent rates: the Drive mode stabilizes the closed state connected to the open state, whereas the Park mode stabilizes the disconnected closed state. Intermediate calcium concentrations are shown to depress the transition rate from Drive to Park mode, restricting frequent Park mode activity to low or high calcium levels.
What carries the argument
Hierarchical Markov chain models with missed-event correction integrated directly into the Bayesian likelihood function for parameter inference and model selection.
Load-bearing premise
The specific functional form and hierarchical structure used to correct for missed events in the likelihood function accurately represent the underlying true gating kinetics without introducing new biases.
What would settle it
High-temporal-resolution recordings that detect all short events, when analyzed without correction, should yield parameters matching those from the corrected lower-resolution data if the method is accurate.
Figures
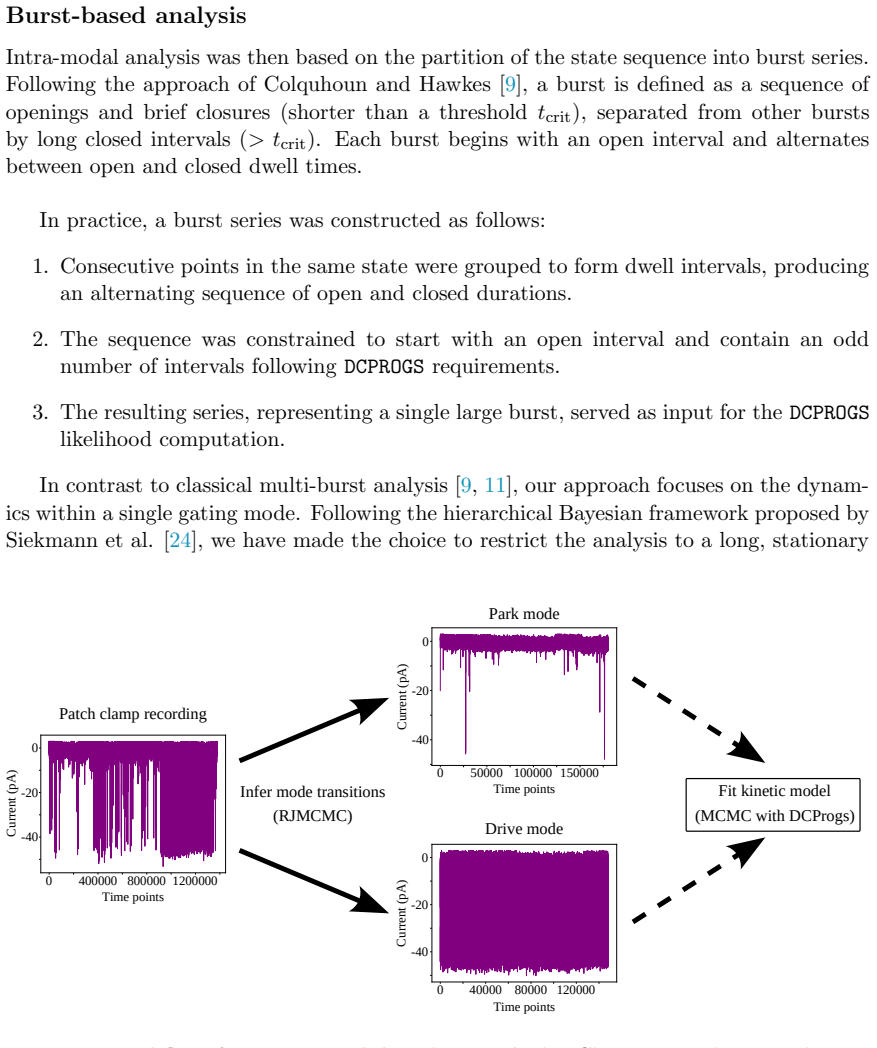

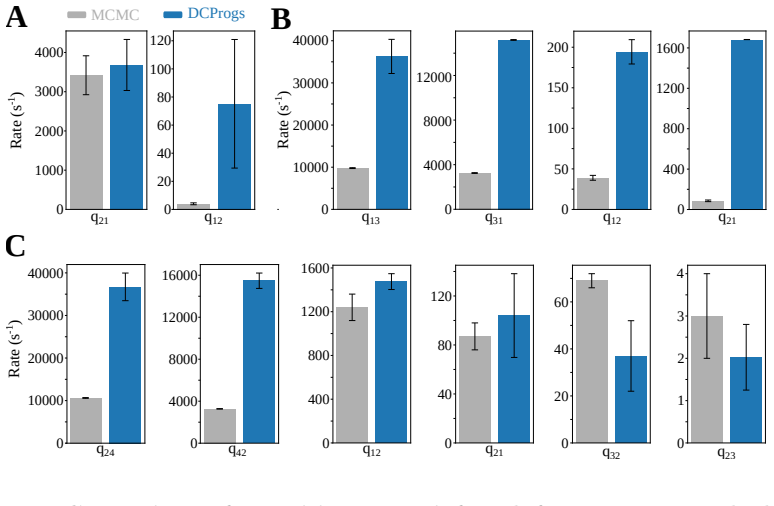
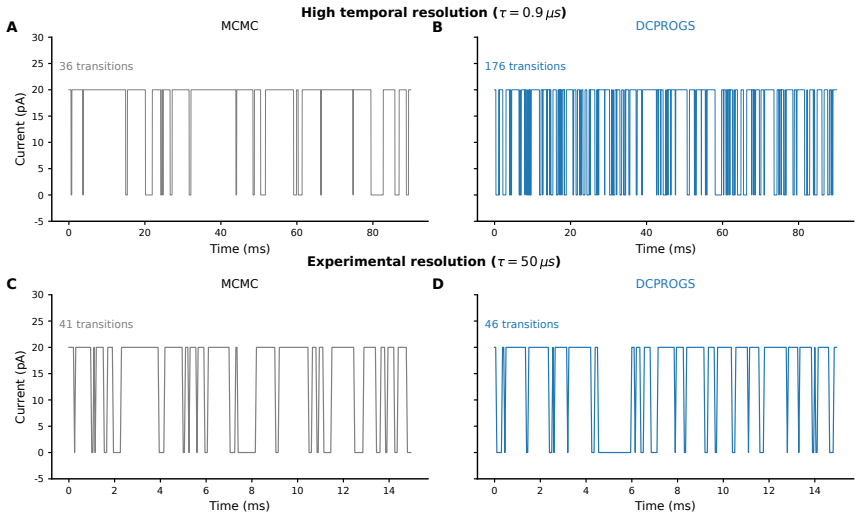
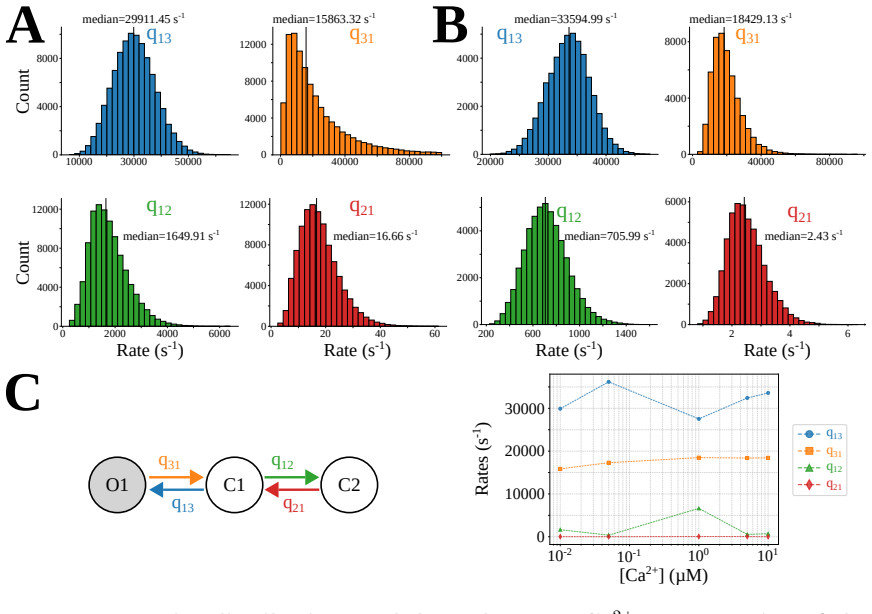



read the original abstract
The Inositol 1,4,5-trisphosphate receptor channel (IP 3 R) is an important calcium channel involved in calcium-induced calcium release, playing a prominent role in intracellular calcium signaling. However, accurately characterizing its gating behavior remains a challenge, particularly due to the temporal resolution of patch clamp techniques that is not large enough to detect all short-lived events. This limitation can significantly bias the inference of kinetic models describing the receptor activity. To address this issue, we focused on the quantitative analysis of IP 3 R gating behavior using patch clamp data, with particular attention to missed events. We modeled IP 3 R channel gating using Hierarchical Markov chains and used a Bayesian approach that integrates missed event correction directly into the likelihood function, enabling more accurate parameter inference and model evaluation. We show that accounting for missed events deeply clarifies the multi-modal model that emerges from model selection. In this new model, the Park and Drive modes both consist of the same 3-state Markov model, with mode-dependent kinetic parameters: the Drive mode stabilizes the closed state directly connected to the open one, whereas the Park mode stabilizes the other closed state, that is not connected to the open one. Intermediate Ca 2+ concentrations are found to strongly depress the Drive to Park transition rate, so that the IP 3 R channel undergoes frequent transitions to the Park mode only for __ 50 nM or micromolar Ca 2+ concentrations. Overall, our approach provides a refined perspective on IP 3 R channel modeling and highlights the critical importance of accounting for missed events upon model selection based on single-channel recordings.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a Bayesian framework for inferring hierarchical Markov models of IP3R single-channel gating from patch-clamp recordings. It integrates an analytic correction for missed brief events (arising from finite temporal resolution) directly into the likelihood, then performs model selection across candidate topologies and parameterizations. The central result is that this correction yields a clarified multimodal description in which both the Park and Drive modes are realized by the same 3-state Markov chain, but with mode-specific rate constants: the Drive mode stabilizes the closed state adjacent to the open state while the Park mode stabilizes the disconnected closed state; in addition, intermediate Ca2+ concentrations strongly suppress the Drive-to-Park transition rate, restricting frequent Park-mode visits to sub-50 nM or micromolar Ca2+ regimes.
Significance. If the missed-event correction is shown to be unbiased, the work supplies a concrete, falsifiable 3-state mechanistic picture of IP3R modal gating together with a quantitative Ca2+ dependence that can be tested in future experiments. The explicit embedding of the correction inside the likelihood and the use of Bayesian model selection constitute a methodological advance that could be adopted for other channels where brief sojourns are routinely missed. The paper thereby strengthens the link between experimental bandwidth limitations and inferred kinetic schemes.
major comments (2)
- [Methods] Methods (likelihood construction): the functional form chosen for the missed-event probability (typically an approximation based on filter dead-time and noise) is inserted into the likelihood without accompanying simulation recovery tests on synthetic data generated from the final 3-state model; because the abstract states that the multimodal clarification appears only after the correction, the absence of such ground-truth validation leaves open the possibility that the reported topology and mode-specific stabilizations are artifacts of the correction rather than features of the data.
- [Results] Results (model selection): no table or figure directly compares posterior model probabilities or Bayes factors obtained with versus without the missed-event term; the claim that accounting for missed events 'deeply clarifies' the 3-state description therefore rests on an implicit before-after contrast that is not quantified, weakening the assertion that the correction is the decisive factor.
minor comments (3)
- [Abstract] Abstract: the phrase 'for __ 50 nM' is a clear typesetting placeholder and should be replaced by the intended inequality (e.g., '< 50 nM').
- [Throughout] Notation: ensure that state labels (C1, C2, O) and rate symbols are defined once and used consistently in both text and any supplementary equations.
- [Figures] Figures: if dwell-time histograms or posterior predictive checks are shown, they should be accompanied by the corresponding uncorrected versions to allow visual assessment of the correction's effect.
Simulated Author's Rebuttal
We thank the referee for their constructive comments on our manuscript. We address each major point below and describe the revisions we will make to strengthen the validation of the missed-event correction and the model-selection results.
read point-by-point responses
-
Referee: [Methods] Methods (likelihood construction): the functional form chosen for the missed-event probability (typically an approximation based on filter dead-time and noise) is inserted into the likelihood without accompanying simulation recovery tests on synthetic data generated from the final 3-state model; because the abstract states that the multimodal clarification appears only after the correction, the absence of such ground-truth validation leaves open the possibility that the reported topology and mode-specific stabilizations are artifacts of the correction rather than features of the data.
Authors: We agree that explicit recovery tests on synthetic data are needed to rule out artifacts. We will add a dedicated subsection to the Methods describing simulation-based validation: synthetic single-channel records will be generated from the final 3-state hierarchical Markov model using the inferred mode-specific rate constants, realistic filter dead-time and noise will be imposed to produce missed events, and the Bayesian procedure will then be reapplied to recover the original topology, mode assignments, and parameter values. These results will be presented in a new supplementary figure. revision: yes
-
Referee: [Results] Results (model selection): no table or figure directly compares posterior model probabilities or Bayes factors obtained with versus without the missed-event term; the claim that accounting for missed events 'deeply clarifies' the 3-state description therefore rests on an implicit before-after contrast that is not quantified, weakening the assertion that the correction is the decisive factor.
Authors: We acknowledge that an explicit side-by-side comparison is required to quantify the effect of the missed-event term. We will add a new table (or supplementary figure) that reports the posterior model probabilities and Bayes factors for the candidate topologies both with and without the missed-event probability in the likelihood. This will directly demonstrate the change in model preference and the stabilization of the 3-state description once the correction is included. revision: yes
Circularity Check
No significant circularity; model emerges from data-driven selection on corrected likelihood
full rationale
The paper integrates a missed-event correction into the likelihood for hierarchical Markov models and performs Bayesian model selection on patch-clamp data. The reported 3-state structure per mode, mode-specific stabilizations, and Ca2+-dependent transition suppression are outputs of that inference and comparison process. No equation or step reduces a claimed prediction or model feature to a quantity defined solely by the fitted parameters themselves, nor does any load-bearing premise collapse to a self-citation or ansatz smuggled from prior author work. The derivation remains self-contained against the experimental recordings and standard single-channel analysis techniques.
Axiom & Free-Parameter Ledger
free parameters (1)
- mode-dependent transition rates
axioms (2)
- standard math Channel gating obeys the Markov property (memoryless transitions between discrete states)
- domain assumption Missed events arise from finite temporal resolution and can be corrected by integrating over unobserved short sojourns in the likelihood
Reference graph
Works this paper leans on
-
[1]
K. Ball, T. G. Kurtz, L. Popovic, and G. Rempala. Asymptotic analysis of multiscale approximations to reaction networks. The Annals of Applied Probability , 16(4):1925– 1961, Nov. 2006. Publisher: Institute of Mathematical Statistics
work page 1925
-
[2]
M. J. Berridge. Calcium microdomains: Organization and function. Cell Calcium , 40(5):405–412, Nov. 2006
work page 2006
-
[3]
L. Bezprozvanny, J. Watras, and B. E. Ehrlich. Bell-shaped calcium-response curves of lns(l,4,5)P3- and calcium-gated channels from endoplasmic reticulum of cerebellum. Nature, 351(6329):751–754, June 1991. Publisher: Nature Publishing Group
work page 1991
-
[4]
A. L. Blatz and K. L. Magleby. Correcting single channel data for missed events. Biophysical Journal, 49(6):1183–1191, June 1986. Publisher: Elsevier
work page 1986
-
[5]
V. Burzomato, M. Beato, P. J. Groot-Kormelink, D. Colquhoun, and L. G. Sivilotti. Single-Channel Behavior of Heteromeric α1β Glycine Receptors: An Attempt to De- tect a Conformational Change before the Channel Opens. Journal of Neuroscience , 24(48):10924–10940, Dec. 2004. Publisher: Society for Neuroscience Section: Cellu- lar/Molecular
work page 2004
-
[6]
P. Cao, X. Tan, G. Donovan, M. J. Sanderson, and J. Sneyd. A Deterministic Model Predicts the Properties of Stochastic Calcium Oscillations in Airway Smooth Muscle Cells. PLOS Computational Biology , 10(8):e1003783, Aug. 2014. Publisher: Public Library of Science. 23
work page 2014
-
[7]
D. Colquhoun and A. G. Hawkes. On the stochastic properties of bursts of single ion channel openings and of clusters of bursts. Philosophical Transactions of the Royal Society of London. B, Biological Sciences , 300(1098):1–59, Dec. 1982
work page 1982
-
[8]
D. Colquhoun and A. G. Hawkes. Stochastic properties of ion channel openings and bursts in a membrane patch that contains two channels: evidence concerning the num- ber of channels present when a record containing only single openings is observed. Pro- ceedings of the Royal Society of London. Series B: Biological Sciences , 240(1299):453– 477, June 1990. ...
work page 1990
-
[9]
D. Colquhoun, A. G. Hawkes, and K. Srodzinski. Joint Distributions of Apparent Open and Shut Times of Single-Ion Channels and Maximum Likelihood Fitting of Mecha- nisms. Philosophical Transactions: Mathematical, Physical and Engineering Sciences , 354(1718):2555–2590, 1996. Publisher: The Royal Society
work page 1996
-
[10]
G. W. De Young and J. Keizer. A single-pool inositol 1,4,5-trisphosphate-receptor- based model for agonist-stimulated oscillations in Ca2+ concentration. Proceedings of the National Academy of Sciences , 89(20):9895–9899, Oct. 1992. Publisher: Proceed- ings of the National Academy of Sciences
work page 1992
-
[11]
M. Epstein, B. Calderhead, M. A. Girolami, and L. G. Sivilotti. Bayesian Statistical Inference in Ion-Channel Models with Exact Missed Event Correction. Biophysical Journal, 111(2):333–348, July 2016
work page 2016
-
[12]
E. Gin, M. Falcke, L. Wagner, D. Yule, and J. Sneyd. Markov chain Monte Carlo fitting of single-channel data from inositol trisphosphate receptors. Journal of theoretical biology, 257:460–74, Apr. 2009
work page 2009
-
[13]
P. J. GREEN. Reversible jump Markov chain Monte Carlo computation and Bayesian model determination. Biometrika, 82(4):711–732, Dec. 1995
work page 1995
-
[14]
O. P. Hamill, A. Marty, E. Neher, B. Sakmann, and F. J. Sigworth. Improved patch- clamp techniques for high-resolution current recording from cells and cell-free mem- brane patches. Pfl¨ ugers Archiv, 391(2):85–100, Aug. 1981
work page 1981
- [15]
-
[16]
A. G. Hawkes, A. Jalali, and D. Colquhoun. The distributions of the apparent open times and shut times in a single channel record when brief events cannot be detected. Philosophical Transactions of the Royal Society of London, Series A: Physical and Engineering Sciences, 332(1627):511–538, Sept. 1990
work page 1990
-
[17]
K. E. Hines. A Primer on Bayesian Inference for Biophysical Systems. Biophysical Journal, 108(9):2103–2113, May 2015. 24
work page 2015
-
[18]
R. Horn. Estimating the number of channels in patch recordings. The Journal of Gen- eral Physiology, 101(3):453–458, Mar. 1991. Publisher: Rockefeller University Press
work page 1991
-
[19]
L. M. Ionescu, C. White, K.-H. Cheung, J. Shuai, I. Parker, J. E. Pearson, J. K. Foskett, and D.-O. D. Mak. Mode switching is the major mechanism of ligand regulation of insp3 receptor calcium release channels. The Journal of General Physiology , 130:631 – 645, 2007
work page 2007
-
[20]
R. E. Kass and A. E. Raftery. Bayes factors. Journal of the American Statistical Association, 90(430):773–795, 1995
work page 1995
-
[21]
D.-O. D. Mak and J. K. Foskett. Inositol 1,4,5-trisphosphate receptors in the endo- plasmic reticulum: A single-channel point of view. Cell Calcium , 58(1):67–78, July 2015
work page 2015
- [22]
-
[23]
E. Oikonomou, Y. Juli, R. R. Kolan, L. Kern, T. Gruber, C. Alzheimer, P. Krauss, A. Maier, and T. Huth. A deep learning approach to real-time Markov modeling of ion channel gating. Communications Chemistry, 7(1):1–16, Nov. 2024. Publisher: Nature Publishing Group
work page 2024
-
[24]
I. Siekmann, M. Fackrell, E. J. Crampin, and P. Taylor. Modelling modal gating of ion channels with hierarchical Markov models. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences , 472(2192):20160122, Aug. 2016
work page 2016
-
[25]
I. Siekmann, J. Sneyd, and E. Crampin. MCMC Can Detect Nonidentifiable Models. Biophysical Journal, 103(11):2275–2286, Dec. 2012
work page 2012
-
[26]
I. Siekmann, J. Sneyd, and E. J. Crampin. Statistical analysis of modal gating in ion channels. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 470(2166):20140030, June 2014. Publisher: Royal Society
work page 2014
-
[27]
I. Siekmann, L. E. Wagner, D. Yule, E. J. Crampin, and J. Sneyd. A Kinetic Model for Type I and II IP3R Accounting for Mode Changes. Biophysical Journal, 103(4):658– 668, Aug. 2012
work page 2012
- [28]
- [29]
-
[30]
L. E. Wagner II and D. I. Yule. Differential regulation of the InsP3 receptor type-1 and -2 single channel properties by InsP3, Ca2+ and ATP. The Journal of Physiology , 590(14):3245–3259, 2012. 26
work page 2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.