A Bias-Corrected Weighted Logistic Model for Gene Regulatory Networks: Functional Equivalence with the Product-of-Logistics and Comparison with Weighted-Sum Formulations
Pith reviewed 2026-06-28 03:52 UTC · model grok-4.3
The pith
A single-sigmoid model with in-degree-dependent bias recovers the equilibria of the product-of-logistics gene network model
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The bias-corrected weighted-logistic model uses the regulatory function σ(λ S_i + λ b_i) with b_i = -λ^{-1} log(2^{m_i}-1) to recover the critical-point value 1/2^{m_i} and the canonical equilibrium x_i^* = κ_i/(γ_i 2^{m_i}) of the product-of-logistics model, and is the unique such single-sigmoid correction. It yields an exact algebraic identity for the discrepancy with the product form, a slope ratio of 2-2^{1-m_i} at the critical point, and the Jacobian relation J^{bcw}=D(J^{prod}+Γ)-Γ with D=diag(2-2^{1-m_i}) at shared equilibria. In the regime γ_i>κ_i λ m_i/4 both systems converge globally and exponentially at rate α=min_i(γ_i-κ_i λ m_i/4).
What carries the argument
The state-independent combinatorial bias b_i depending only on in-degree m_i and steepness λ, inserted inside a single logistic sigmoid of the weighted regulator sum to enforce matching at the critical point and equilibrium.
If this is right
- The bcw and product-of-logistics models share the same equilibria under self-consistency.
- In the contractive regime γ_i>κ_i λ m_i/4 both models converge globally and exponentially to the shared equilibrium at the same explicit rate.
- At any shared equilibrium the Jacobians are related by the diagonal scaling D=diag(2-2^{1-m_i}), producing a stability dichotomy.
- The bcw form undergoes Hopf bifurcation at 2/3 of the product threshold in a two-gene negative-feedback motif and sustains limit cycles where the product form is linearly stable.
Where Pith is reading between the lines
- The explicit discrepancy identity as a sum over regulator subsets could be used to quantify approximation error in large networks without simulating both models.
- The mean-field reading of the bias as a zero-order log-sum-exp approximation points to possible higher-order corrections that might reduce curvature mismatch for m_i>=2.
- The no-go result on global equivalence by any state-independent constant suggests that state-dependent or higher-order terms would be required for exact trajectory matching.
- The eigenvalue amplification by D in the Jacobian relation implies that local stability margins differ even when equilibria coincide, which could affect robustness predictions in network design.
Load-bearing premise
The regulatory input for each gene can be expressed as a single sigmoid applied to a linear weighted sum of regulator states plus a bias that depends only on the in-degree and steepness parameter.
What would settle it
Measuring steady-state expression levels in a gene network whose in-degrees and parameters place it in the contractive regime, then checking whether they equal κ_i/(γ_i 2^{m_i}) under the bias-corrected model, would confirm or refute the shared-equilibrium claim.
Figures

read the original abstract
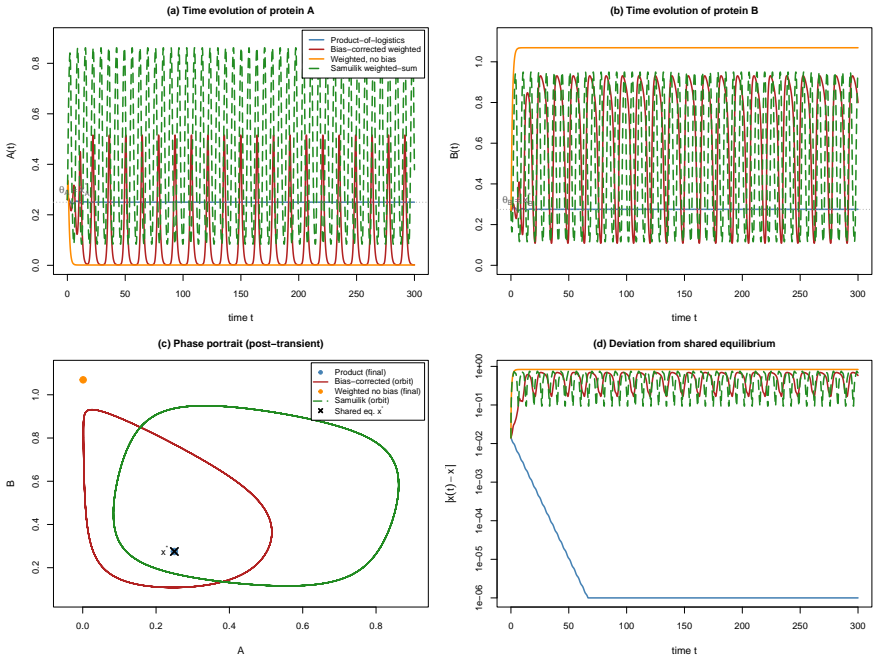
We introduce a bias-corrected weighted-logistic (bcw) formulation for ODE models of gene regulatory networks. Each gene's regulatory function is a single sigmoid $\sigma(\lambda S_i+\lambda b_i)$ of the signed weighted regulator sum $S_i$, with a combinatorial bias $b_i=-\lambda^{-1}\log(2^{m_i}-1)$ depending only on the in-degree $m_i$ and the steepness $\lambda$. We prove this is the unique single-sigmoid correction recovering the critical-point value $1/2^{m_i}$ of the product-of-logistics model and sharing its canonical equilibrium $x_i^*=\kappa_i/(\gamma_i 2^{m_i})$ under self-consistency. The bcw system inherits global $C^\infty$ well-posedness, an explicit Lipschitz bound, a positively invariant box, and strictly positive basal output. We derive an exact algebraic identity for the discrepancy $f^{prod}-f^{bcw}$ as a sum over proper non-empty regulator subsets; a slope ratio $2-2^{1-m_i}$ at the critical point; a curvature mismatch breaking the symmetry $\sigma''=0$ for $m_i\ge2$; a no-go result excluding global equivalence by any state-independent constant; and a mean-field reading of the bias as a zero-order log-sum-exp approximation. At a shared equilibrium the Jacobians satisfy $J^{bcw}=D(J^{prod}+\Gamma)-\Gamma$ with $D=\mathrm{diag}(2-2^{1-m_i})$, giving a stability dichotomy. In the contractive regime $\gamma_i>\kappa_i\lambda m_i/4$ both systems converge globally and exponentially to the shared equilibrium at rate $\alpha=\min_i(\gamma_i-\kappa_i\lambda m_i/4)$. A two-gene self-activating negative-feedback motif illustrates the theory: the bcw form undergoes Hopf bifurcation at $2/3$ of the product threshold and sustains limit cycles where the product is linearly stable, with eigenvalue amplification factor $D=3/2$.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces a bias-corrected weighted-logistic (bcw) formulation for ODE models of gene regulatory networks. Each regulatory function is a single sigmoid σ(λ S_i + λ b_i) with combinatorial bias b_i = −λ^{-1} log(2^{m_i}−1) depending only on in-degree m_i and steepness λ. It proves this bias is the unique single-sigmoid choice recovering the product-of-logistics critical-point value 1/2^{m_i} and the canonical equilibrium x_i^* = κ_i/(γ_i 2^{m_i}) under self-consistency. The manuscript derives an exact algebraic identity for the discrepancy f^{prod}−f^{bcw}, a slope ratio 2−2^{1−m_i}, a curvature mismatch, a no-go result excluding global equivalence by any state-independent constant, and the Jacobian relation J^{bcw}=D(J^{prod}+Γ)−Γ at equilibrium with D=diag(2−2^{1−m_i}). It establishes global convergence in the contractive regime γ_i > κ_i λ m_i/4 and illustrates via a two-gene self-activating negative-feedback motif that bcw undergoes Hopf bifurcation at 2/3 the product threshold while sustaining limit cycles where the product model is linearly stable.
Significance. If the algebraic identities, uniqueness argument, and Jacobian relation hold, the bcw model supplies a computationally simpler single-sigmoid alternative to product-of-logistics formulations while exactly preserving equilibria and supplying an explicit global exponential convergence rate α=min_i(γ_i−κ_i λ m_i/4). The explicit bias choice, the mean-field log-sum-exp interpretation, and the stability dichotomy via the diagonal factor D are concrete strengths that could aid analysis of larger networks. The two-gene example demonstrates that local linearization differences can produce qualitatively distinct global dynamics, which is a useful cautionary result.
major comments (2)
- [bcw formulation and uniqueness proof] The uniqueness claim for the bias (recovering both the critical-point value and the canonical equilibrium) is load-bearing for the central contribution; the derivation from σ(λ b_i)=2^{-m_i} is stated, but the manuscript should explicitly verify that the self-consistency condition x_i^*=κ_i/(γ_i 2^{m_i}) is satisfied simultaneously without additional assumptions on the production and degradation rates.
- [Jacobian and stability analysis] The Jacobian relation J^{bcw}=D(J^{prod}+Γ)−Γ with D=diag(2−2^{1−m_i}) is used to obtain the stability dichotomy and the contractive-regime convergence rate; this relation is stated to hold at the shared equilibrium, but the paper must confirm whether the factor D preserves the global Lipschitz bound and positive invariance already claimed for the bcw system or introduces any additional restrictions.
minor comments (2)
- [discrepancy identity] The algebraic identity for the discrepancy f^{prod}−f^{bcw} as a sum over proper non-empty regulator subsets is a useful explicit result; it would benefit from a short worked example for m_i=2 or m_i=3 to illustrate the terms.
- [model definition] Notation for the steepness parameter λ and the in-degree m_i is consistent, but the manuscript should clarify early whether λ is assumed identical across all nodes or allowed to vary.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive comments. We address each major comment below.
read point-by-point responses
-
Referee: [bcw formulation and uniqueness proof] The uniqueness claim for the bias (recovering both the critical-point value and the canonical equilibrium) is load-bearing for the central contribution; the derivation from σ(λ b_i)=2^{-m_i} is stated, but the manuscript should explicitly verify that the self-consistency condition x_i^*=κ_i/(γ_i 2^{m_i}) is satisfied simultaneously without additional assumptions on the production and degradation rates.
Authors: We agree an explicit verification is useful. With b_i chosen so that σ(λ b_i)=2^{-m_i}, the canonical equilibrium x_i^*=κ_i/(γ_i 2^{m_i}) is defined precisely so that the weighted sum S_i(x^*) equals the point at which the product model equals 1/2^{m_i}. Substituting into the bcw sigmoid then yields f^{bcw}(x^*)=γ_i x_i^*/κ_i by direct algebra, for any positive κ_i, γ_i. In revision we add a short calculation immediately after the bias derivation confirming this holds under self-consistency alone. revision: yes
-
Referee: [Jacobian and stability analysis] The Jacobian relation J^{bcw}=D(J^{prod}+Γ)−Γ with D=diag(2−2^{1−m_i}) is used to obtain the stability dichotomy and the contractive-regime convergence rate; this relation is stated to hold at the shared equilibrium, but the paper must confirm whether the factor D preserves the global Lipschitz bound and positive invariance already claimed for the bcw system or introduces any additional restrictions.
Authors: The identity J^{bcw}=D(J^{prod}+Γ)−Γ holds only at the shared equilibrium and does not modify the bcw vector field. The global Lipschitz bound, positive invariance of the unit box, and C^∞ well-posedness are proved directly for the bcw ODE (Section 3) and are independent of D. We will insert one clarifying sentence in the stability section stating that these global properties remain unchanged. revision: yes
Circularity Check
No significant circularity identified
full rationale
The paper explicitly introduces the bias b_i = −λ^{-1} log(2^{m_i}−1) to enforce σ(λ b_i) = 2^{-m_i} by algebraic solution of the logistic sigmoid equation, then proves uniqueness of this choice for matching the product model's critical-point value at S_i=0 and the resulting equilibrium under self-consistency. This is a deliberate, transparent construction rather than a hidden reduction; the Jacobian relation J^{bcw}=D(J^{prod}+Γ)−Γ and other identities follow directly from substituting the stated functional forms. No load-bearing self-citation, no parameter fitted to data then relabeled as prediction, and no ansatz or uniqueness theorem imported from prior author work. The paper acknowledges the no-go result for global equivalence. The derivation chain is self-contained and externally verifiable from the given assumptions.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The regulatory function is a composition of the standard logistic sigmoid with a linear weighted sum plus bias.
- domain assumption The product-of-logistics model serves as the baseline whose critical value and equilibrium must be recovered.
Reference graph
Works this paper leans on
-
[1]
Abramowitz and I
M. Abramowitz and I. A. Stegun.Handbook of Mathematical Functions with Formulas, Graphs, and Mathematical Tables. Dover, 1965
1965
-
[2]
Boyd and L
S. Boyd and L. Vandenberghe.Convex Optimization. Cambridge University Press, 2004. 37
2004
-
[3]
M. J. Wainwright and M. I. Jordan. Graphical models, exponential families, and varia- tional inference.Foundations and Trends in Machine Learning, 1(1–2):1–305, 2008
2008
-
[4]
Glass and S
L. Glass and S. A. Kauffman. The logical analysis of continuous, non-linear biochemical control networks.Journal of Theoretical Biology, 39(1):103–129, 1973
1973
-
[5]
M. W. Hirsch and H. L. Smith. Monotone dynamical systems.Handbook of Differential Equations: Ordinary Differential Equations, 2:239–357, 2005
2005
-
[6]
Vohradsky
J. Vohradsky. Neural network model of gene expression.The FASEB Journal, 15(3):846– 854, 2001
2001
-
[7]
I. Belgacem. Exploring logistic functions as robust alternatives to Hill functions in genetic network modeling. arXiv preprint2512.14325, 2025. Available athttps://arxiv.org/ abs/2512.14325
Pith/arXiv arXiv 2025
-
[8]
I. Belgacem. Logistic gene regulatory networks: prevention of expression shutdown, and numerical stability beyond Hill functions. arXiv preprint2605.01056, 2026. Available at https://arxiv.org/abs/2605.01056
Pith/arXiv arXiv 2026
-
[9]
I. Belgacem. Beyond linear additive and Hill functions: a general logistic reformulation of delay-coupled gene regulatory networks with equilibrium analysis, Hopf bifurcation, and Lipschitz stability. arXiv preprint2604.26810, 2026. Available athttps://arxiv.org/ abs/2604.26810
Pith/arXiv arXiv 2026
-
[10]
I. Belgacem. Sustained limit cycles in the logistic two-gene genetic oscillator: a delay- driven Hopf bifurcation. arXiv preprint2605.23722, 2026. Available athttps://arxiv. org/abs/2605.23722
Pith/arXiv arXiv 2026
-
[11]
I. Belgacem, S. Casagranda, E. Grac, D. Ropers, and J.-L. Gouz´ e. Reduction and stability analysis of a transcription-translation model of RNA polymerase.Bulletin of Mathematical Biology, 80:294–318, 2018. DOI:10.1007/s11538-017-0372-4
-
[12]
I. Belgacem and J.-L. Gouz´ e. Mathematical study of the global dynamics of a concave gene-expression model. In22nd Mediterranean Conference on Control and Automation (MED’14), Palermo, Italy, pages 1341–1346. IEEE, 2014. DOI:10.1109/MED.2014. 6961562
-
[13]
I. Belgacem, R. Edwards, and ´E. Farcot. Computer-aided analysis of high-dimensional Glass networks: periodicity, chaos, and bifurcations in a ring circuit.Chaos: An Interdis- ciplinary Journal of Nonlinear Science, 35(5), May 2025. DOI:10.1063/5.0243955
-
[14]
I. Belgacem, J.-L. Gouz´ e, and R. Edwards. Control of negative feedback loops in genetic networks. InProceedings of the 59th IEEE Conference on Decision and Control (CDC), Jeju Island, Republic of Korea, pages 5098–5105. IEEE, 2020. DOI:10.1109/CDC42340. 2020.9304088
-
[15]
L. Chambon, I. Belgacem, and J.-L. Gouz´ e. Qualitative control of undesired oscillations in a genetic negative feedback loop with uncertain measurements.Automatica, 112:108642, February 2020. DOI:10.1016/j.automatica.2019.108642
-
[16]
Farcot, S
E. Farcot, S. Best, R. Edwards, I. Belgacem, X. Xu, and P. Gill. Chaos in a ring circuit. Chaos: An Interdisciplinary Journal of Nonlinear Science, 29(4):043103, 2019. DOI:10. 1063/1.5079941. 38
2019
-
[17]
I. Belgacem, H. Bensalah, B. Cherki, and R. Edwards. The probabilistic convolution regularization of Zeno hybrid systems. In18th European Control Conference (ECC), Naples, Italy, pages 2017–2022. IEEE, June 2019. DOI:10.23919/ECC.2019.8795626
-
[18]
Belgacem, E
I. Belgacem, E. Grac, D. Ropers, and J.-L. Gouz´ e. Stability analysis of a reduced transcription-translation model of RNA polymerase. InIEEE 53rd Conference on De- cision and Control (CDC), Los Angeles, CA, USA, pages 3924–3929. IEEE, December
-
[19]
DOI:10.1109/CDC.2014.7039999
-
[20]
I. Belgacem and J.-L. Gouz´ e. Stability analysis and reduction of gene transcription models. InIEEE 52nd Annual Conference on Decision and Control (CDC), Florence, Italy, pages 2691–2696. IEEE, 2013. DOI:10.1109/CDC.2013.6760289
-
[21]
Belgacem and J.-L
I. Belgacem and J.-L. Gouz´ e. Global stability of enzymatic chain of full reversible Michaelis–Menten reactions.Acta Biotheoretica, 61(3):425–436, 2013. DOI:10.1007/ s10441-013-9195-3
2013
-
[22]
I. Belgacem and J.-L. Gouz´ e. Analysis and reduction of transcription–translation cou- pled models for gene expression. In12th IFAC Symposium on Computer Applications in Biotechnology (CAB), Mumbai, India, pages 42–47. Elsevier, 2013. DOI:10.3182/ 20131216-3-IN-2044.00012
arXiv 2013
-
[23]
I. Belgacem and J.-L. Gouz´ e. Global stability of full open reversible Michaelis–Menten reactions. In8th IFAC Symposium on Advanced Control of Chemical Processes, Singapore, pages 591–596. Elsevier, July 2012. DOI:10.3182/20120710-4-SG-2026.00039
-
[24]
Bintu, N
L. Bintu, N. E. Buchler, H. G. Garcia, U. Gerland, T. Hwa, J. Kondev, and R. Phillips. Transcriptional regulation by the numbers: models.Current Opinion in Genetics & De- velopment, 15(2):116–124, 2005
2005
-
[25]
T. S. Gardner, C. R. Cantor, and J. J. Collins. Construction of a genetic toggle switch in Escherichia coli.Nature, 403(6767):339–342, 2000
2000
-
[26]
M. B. Elowitz and S. Leibler. A synthetic oscillatory network of transcriptional regulators. Nature, 403(6767):335–338, 2000
2000
-
[27]
M. E. Hern´ andez-Garc´ ıa and J. Vel´ azquez-Castro. Corrected Hill function in stochastic gene regulatory networks. arXiv preprint2307.03057, 2023. DOI:10.48550/arXiv. 2307.03057
work page internal anchor Pith review doi:10.48550/arxiv 2023
-
[28]
O. Kozlovska and F. Sadyrbaev. Models of genetic networks with given properties.WSEAS Transactions on Computer Research, 10:43–49, 2022. DOI:10.37394/232018.2022.10.6
-
[29]
O. Kozlovska and I. Samuilik. Quasi-periodic solutions for a three-dimensional system in gene regulatory network.WSEAS Transactions on Systems, 22:727–733, 2023. DOI:10. 37394/23202.2023.22.73
arXiv 2023
-
[30]
F. Sadyrbaev, I. Samuilik, and V. Sengileyev. On modelling of genetic regulatory networks. WSEAS Transactions on Electronics, 12:73–80, 2021. DOI:10.37394/232017.2021.12. 10
-
[31]
Sadyrbaev, V
F. Sadyrbaev, V. Sengileyev, and A. Silvans. On coexistence of inhibition and activation in genetic regulatory networks.AIP Conference Proceedings, 2849:120004, 2023. DOI:10. 1063/5.0162579. 39
2023
-
[32]
I. Samuilik, F. Sadyrbaev, and S. Atslega. Mathematical modelling of nonlinear dynamic systems. InEngineering for Rural Development (ERDev), volume 21, pages 172–178, Jelgava, Latvia, 2022. DOI:10.22616/ERDev.2022.21.TF051
-
[33]
I. Samuilik, F. Sadyrbaev, and D. Ogorelova. Mathematical modeling of three-dimensional genetic regulatory networks using logistic and Gompertz functions.WSEAS Transactions on Systems and Control, 17:101–107, 2022. DOI:10.37394/23203.2022.17.12
-
[34]
Santill´ an
M. Santill´ an. On the use of the Hill functions in mathematical models of gene regulatory networks.Mathematical Modelling of Natural Phenomena, 3(2):85–97, 2008. DOI:10. 1051/mmnp:2008056
2008
-
[35]
S. S. Somathilaka, S. Balasubramaniam, D. P. Martins, and X. Li. Revealing gene regulation-based neural network computing in bacteria.Biophysical Reports, 3(3):100118,
-
[36]
DOI:10.1016/j.bpr.2023.100118
-
[37]
S. Vinoth, S. L. Kingston, S. Srinivasan, T. Kapitaniak, and A. Venkatesan. Extreme events in gene regulatory networks with time-delays.Scientific Reports, 15:13064, 2025. DOI:10.1038/s41598-025-97268-w. 40 0 50 100 150 200 250 300 0.0 0.2 0.4 0.6 0.8 (a) Time evolution of protein A time t A(t) θA = xA * Product−of−logistics Bias−corrected weighted Weight...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.