Numerical Reliability of Logistic Gene Regulatory Network Models: Preventing Expression Shutdown and Robust Integration of Boolean-Derived ODE Systems
Pith reviewed 2026-05-09 14:34 UTC · model grok-4.3
The pith
Logistic functions fix the smoothness, numerical stability, and zero-basal-rate flaws of Hill functions in gene regulatory network models.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
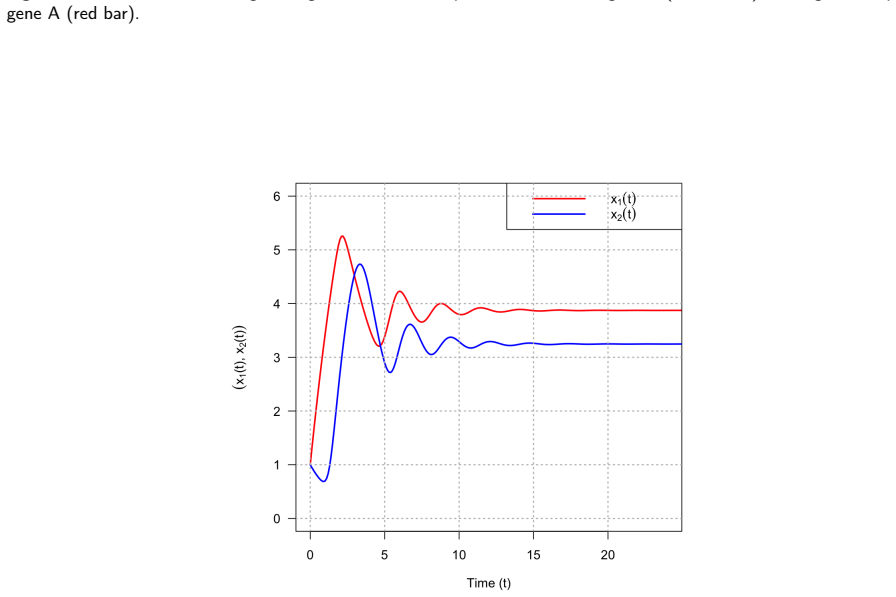
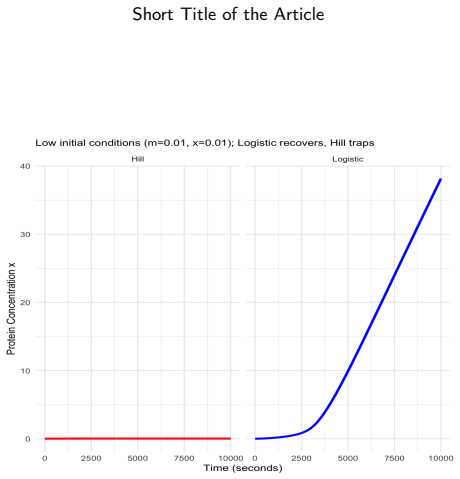
Logistic functions f±, defined to be globally C^∞, real for every real input, and strictly positive at zero, resolve the three structural flaws of Hill functions simultaneously. In a two-gene negative-feedback oscillator, the Routh-Hurwitz criterion shows local asymptotic stability for all positive parameter values and rules out Hopf bifurcation without time delays. For bistable positive autoregulation, saddle-node thresholds are given by explicit transcendental equations; with E. coli parameters the logistic basal rate produces off-state escape in approximately 44 minutes while the Hill model remains trapped indefinitely. The product-of-logistics De Morgan formalism translates Boolean logic
What carries the argument
The logistic activation functions f± that replace Hill functions, guaranteeing global smoothness, real arithmetic, and positive basal production rate.
Load-bearing premise
That logistic functions with positive basal production represent the underlying biology at least as accurately as Hill functions for the gene circuits and parameter ranges considered.
What would settle it
A laboratory measurement showing that a bistable positive-autoregulation circuit remains permanently off under conditions where the logistic model predicts escape within one hour, or a simulation run in which a Hill model with non-integer exponent produces no complex values or warnings.
Figures





read the original abstract
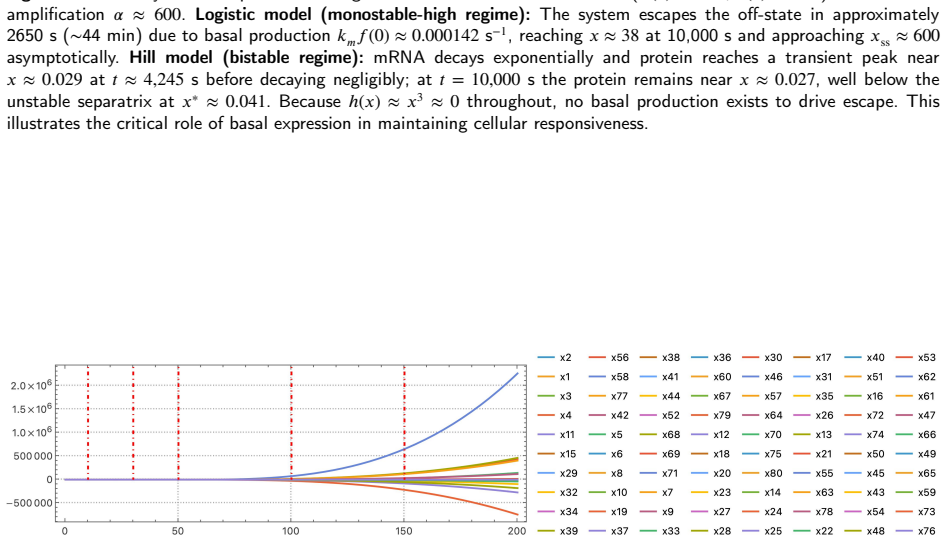
Gene regulatory networks are routinely translated from Boolean update rules into large continuous ODE systems integrated numerically for attractor identification, sensitivity analysis, and control design. The reliability of that integration depends critically on the sigmoidal kernel representing regulation. This simulation study shows that the Hill function -- the near-universal choice -- is a generically unreliable kernel, while the logistic function is a robust replacement. Two failure modes are demonstrated. First, because the Hill function vanishes at zero input, bistable circuits acquire an absorbing off-state: with experimentally grounded \textit{E. coli} galactose-operon autoregulation parameters, a Hill model stays trapped below the unstable separatrix, whereas the logistic model -- whose basal rate is strictly positive by construction -- escapes in about $44$~minutes through basal production alone, matching an analytical estimate of ${\approx}58$~min. A saddle-node analysis characterises the bistable window via an explicit transcendental equation and identifies the threshold $\lambda\theta=2$ separating monostable from bistable regimes. Second, when the Hill exponent is non-integer -- as in dose-response fits -- the power law $x^n=e^{n\ln x}$ turns complex-valued whenever a solver overshoots into negative concentrations. On an $80$-gene Boolean-derived benchmark with $n\approx3.509$, the Hill solver is silently contaminated by complex values from $t\approx52.64$, yielding smooth but spurious trajectories, whereas the logistic formulation completes $t\in[0,200]$ without a single warning. Because the logistic vector field is globally Lipschitz with explicit constant, we further prove an a priori global-error bound of classical order -- a guarantee structurally unavailable to the Hill formulation.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes logistic functions f^± as replacements for Hill functions in gene regulatory network (GRN) models. It argues that logistics simultaneously resolve three Hill-function flaws for non-integer cooperativity: loss of global C^∞ smoothness, silent complex-valued arithmetic corruption of ODE trajectories, and identically zero basal production that traps bistable models in off-states. The paper establishes local asymptotic stability of a two-gene negative-feedback oscillator for all positive parameters via the Routh-Hurwitz criterion with no Hopf bifurcation possible without delays; derives saddle-node thresholds for bistable positive autoregulation via explicit transcendental equations; reports off-state escape in ≈44 min under biophysically grounded E. coli parameters; translates an 11-gene Traynard cell-cycle Boolean network via product-of-logistics De Morgan formalism; and demonstrates that an 80-gene Boolean-derived ODE system (n=3.509) completes t∈[0,200] without warnings while the Hill formulation enters complex-valued contamination at t≈52.64.
Significance. If the logistic basal rate is biologically appropriate, the work supplies a globally smooth, real-valued, and fully controllable framework for GRN modeling that avoids numerical artifacts and enables control-theoretic methods. The Routh-Hurwitz proof, explicit transcendental saddle-node conditions, and reproducible numerical demonstrations on both small and 80-gene systems constitute clear strengths. The De Morgan threshold placement at positive concentrations is a useful contrast to prior weighted-sum formulations. Significance is tempered by the need for stronger justification that non-zero basal production improves rather than distorts bistable dynamics.
major comments (2)
- [Bistable positive autoregulation] Bistable positive autoregulation section: the claim that logistic f^± 'prevents expression shutdown' rests on the finite mean escape time produced exactly by f(0)>0. The manuscript reports ≈44 min escape under 'biophysically grounded E. coli parameters' but does not list the explicit basal production rate, its literature source, or sensitivity analysis; without these, it is impossible to assess whether the result reflects measured promoter leakiness or post-hoc parameter selection that artifactually destabilizes the off-state.
- [Two-gene negative-feedback oscillator] Oscillator stability section: the Routh-Hurwitz criterion is applied to conclude local asymptotic stability for all positive parameters and no Hopf bifurcation without delays. The characteristic equation and the explicit Routh array entries should be provided so readers can verify the sign conditions independently; their absence makes the 'for all positive parameters' claim difficult to check.
minor comments (2)
- [Introduction / Methods] The explicit functional forms of the logistic activation f^+ and repression f^- (including the precise definition of the cooperativity parameter) are referenced but not displayed in the abstract or early sections; they should appear with equation numbers before any stability or bifurcation analysis.
- [Large-network numerical integration] In the 80-gene numerical example the cooperativity n=3.509 is stated without indicating whether it arises from a fit to data or is chosen for illustration; this detail affects reproducibility of the complex-value failure time t≈52.64.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed comments. We address each major point below and will revise the manuscript to incorporate the requested clarifications and supporting material.
read point-by-point responses
-
Referee: [Bistable positive autoregulation] Bistable positive autoregulation section: the claim that logistic f^± 'prevents expression shutdown' rests on the finite mean escape time produced exactly by f(0)>0. The manuscript reports ≈44 min escape under 'biophysically grounded E. coli parameters' but does not list the explicit basal production rate, its literature source, or sensitivity analysis; without these, it is impossible to assess whether the result reflects measured promoter leakiness or post-hoc parameter selection that artifactually destabilizes the off-state.
Authors: We agree that explicit documentation is required. In the revised manuscript we will state the precise basal production rate employed for the E. coli parameter set, cite the primary literature sources for measured promoter leakiness in E. coli, and add a sensitivity analysis that varies the basal rate over the range of experimentally reported values. This will show that off-state escape times remain on the order of tens of minutes for biophysically plausible leakiness levels. revision: yes
-
Referee: [Two-gene negative-feedback oscillator] Oscillator stability section: the Routh-Hurwitz criterion is applied to conclude local asymptotic stability for all positive parameters and no Hopf bifurcation without delays. The characteristic equation and the explicit Routh array entries should be provided so readers can verify the sign conditions independently; their absence makes the 'for all positive parameters' claim difficult to check.
Authors: We will include the full derivation in the revision. The updated section will present the Jacobian of the two-gene system, the resulting characteristic polynomial, and the complete Routh array with all entries and sign conditions. This will allow direct verification that the Hurwitz criteria are satisfied for every positive parameter combination. revision: yes
Circularity Check
No significant circularity; derivations are self-contained
full rationale
The paper derives local stability for the negative-feedback oscillator directly from the Routh-Hurwitz criterion applied to the logistic ODE system, obtains saddle-node thresholds for positive autoregulation via explicit transcendental equations, and performs direct numerical integration on the 11-gene and 80-gene networks. All results follow from the stated functional forms of f^±, the De Morgan translation rule, and externally supplied biophysical parameters without any fitting of outputs to inputs, self-referential definitions, or load-bearing self-citations. The contrast with Hill functions is presented as a transparent consequence of the basal-rate difference rather than a derived prediction.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math The logistic function is globally C^∞, real-valued for all real arguments, and strictly positive at zero
Forward citations
Cited by 3 Pith papers
-
Sustained Limit Cycles in the Logistic Two-Gene Genetic Oscillator: A Delay-Driven Hopf Bifurcation
Delay-differential extension of the logistic two-gene oscillator undergoes a Hopf bifurcation at explicit critical total delay τ_c with closed-form ω_c, transversality bound, and supercritical amplitude laws, matching...
-
State-Feedback Control of Logistic-Based Gene Regulatory Networks: Closed-Form Lyapunov Certificates, Monostabilization, and Delay-Uniform Stability
Introduces logistic-based GRN control with explicit feedforward-proportional laws, quadratic Lyapunov certificates for global exponential stability, monostabilization budgets, and delay-uniform stability theorems.
-
A Bias-Corrected Weighted Logistic Model for Gene Regulatory Networks: Functional Equivalence with the Product-of-Logistics and Comparison with Weighted-Sum Formulations
Introduces bcw single-sigmoid model for GRNs that recovers product-of-logistics critical value 1/2^{m_i} and shared equilibrium, with Jacobian and stability comparisons.
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.