Immune history shapes recurrent epidemics of antigenically related variants
Pith reviewed 2026-07-02 01:40 UTC · model grok-4.3
The pith
Immune history from past variants creates stable recurrent epidemics whose size peaks at intermediate transmission rates.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
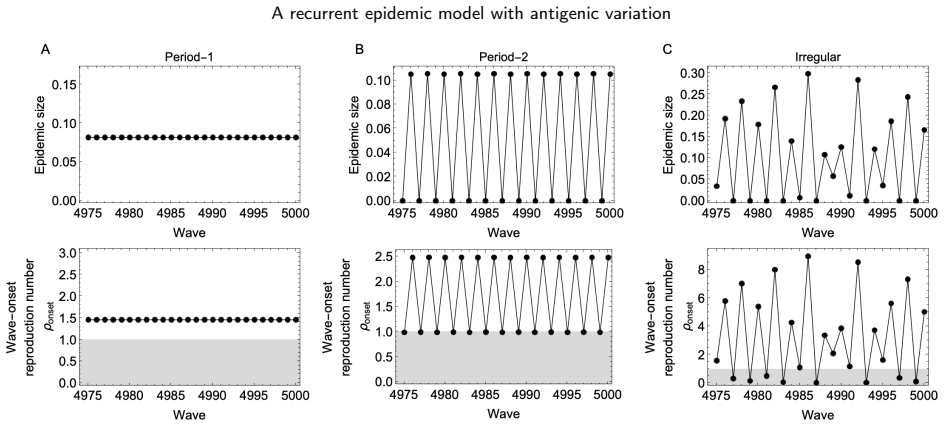
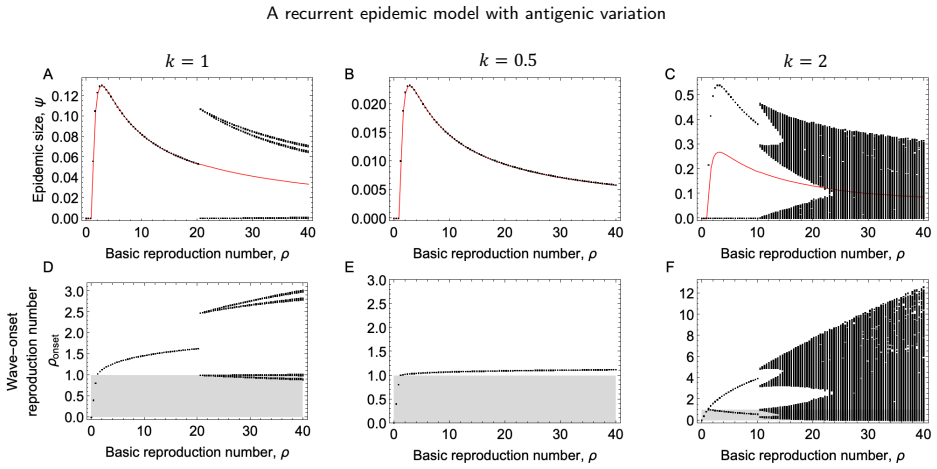
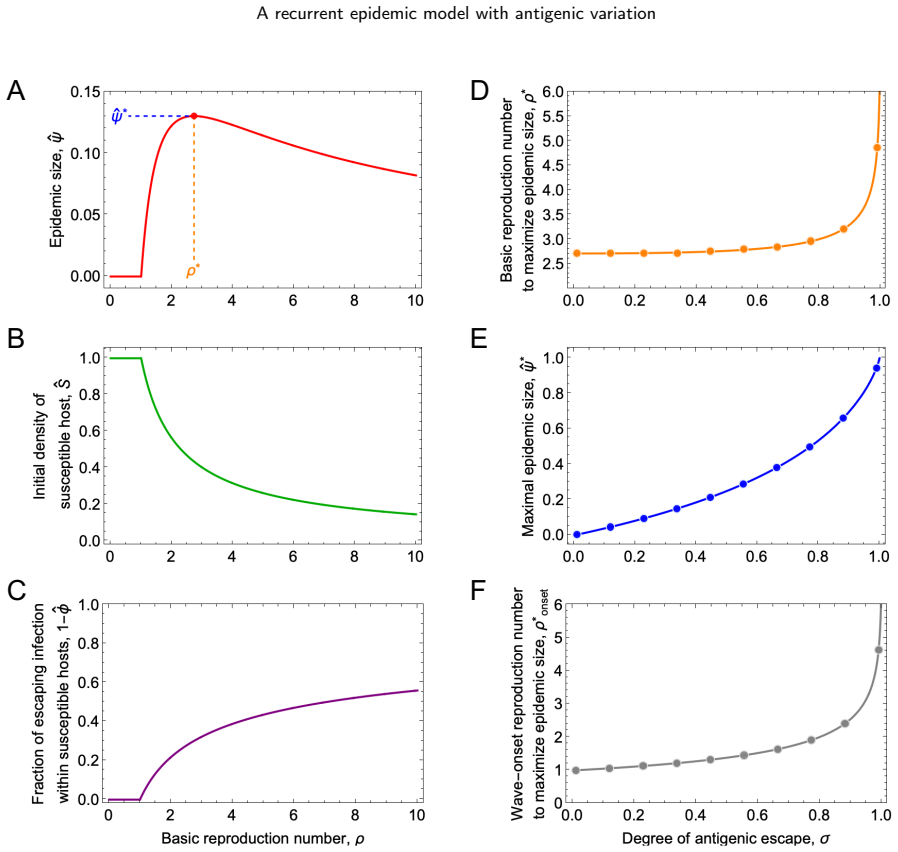
The model reveals that stable, equal-sized recurrent epidemics occur across broad parameter ranges, but can be destabilized when transmission is strong and antigenic escape is limited, leading to period-2 or more complex epidemic dynamics. Epidemic size is maximized at an intermediate basic reproduction number: higher transmissibility boosts immediate infection but also enhances cross-immunity, reducing future susceptibility of the population.
What carries the argument
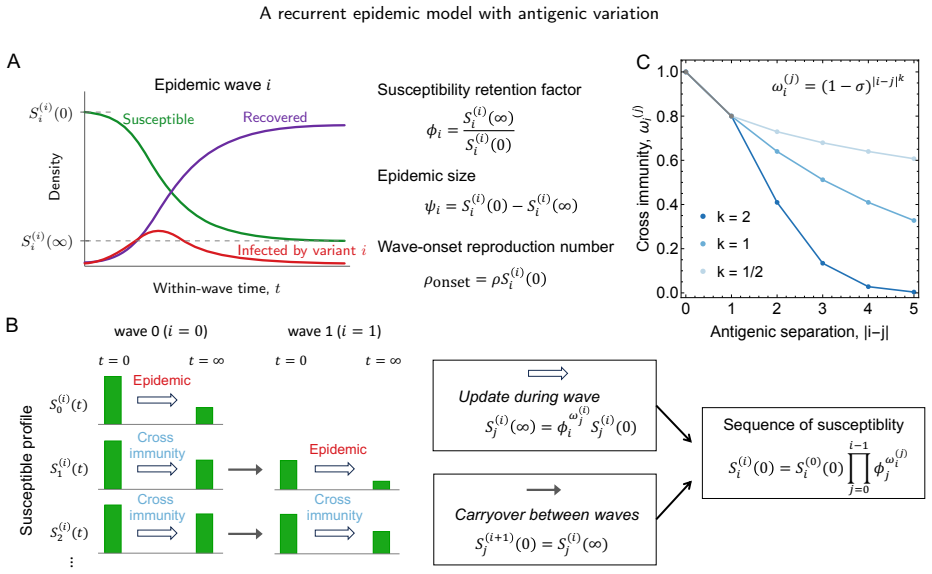
The recurrence map for the population susceptibility to successive variants under the assumption of status-based population immunity.
If this is right
- Stable, equal-sized recurrent epidemics occur across broad parameter ranges.
- The dynamics can destabilize to period-2 or complex when transmission is strong and antigenic escape is limited.
- Epidemic size is maximized at an intermediate basic reproduction number.
- Higher transmissibility increases immediate infection but also cross-immunity reducing future susceptibility.
Where Pith is reading between the lines
- Tracking antigenic similarity between successive variants could improve epidemic forecasting models.
- Reducing transmission rates might sometimes increase the size of future epidemics by limiting cross-immunity development.
- The stability of epidemic cycles depends critically on the rate of antigenic escape.
Load-bearing premise
The model assumes status-based population immunity that allows reduction to a recurrence map for susceptibility to successive variants.
What would settle it
Long-term observation of epidemic sizes in a pathogen with known antigenic distances and transmission rates to check if sizes peak at intermediate R0 or show period-2 oscillations under high transmission.
Figures

read the original abstract
Population immunity carried over from past epidemics of an antigenically variable pathogen influences the epidemic of new variants based on their antigenic similarity to the previous ones. We develop a recurrent SIR model where a population faces sequential, antigenically related variants. The model yields a recurrence map for the population susceptibility to successive variants under the assumption of status-based population immunity. The model reveals that stable, equal-sized recurrent epidemics occur across broad parameter ranges, but can be destabilized when transmission is strong and antigenic escape is limited, leading to period-2 or more, or even more complex epidemic dynamics. Epidemic size is maximized at an intermediate basic reproduction number: higher transmissibility boosts immediate infection but also enhances cross-immunity, reducing future susceptibility of the population. Our results clarify how immune history shapes recurrent epidemics and why success in one wave does not ensure larger future epidemics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a recurrent SIR model for sequential, antigenically related variants. Under the assumption of status-based population immunity, the model yields a recurrence map for the population susceptibility to successive variants. The analysis reveals that stable, equal-sized recurrent epidemics occur across broad parameter ranges, but can be destabilized when transmission is strong and antigenic escape is limited, leading to period-2 or more complex dynamics. Epidemic size is maximized at an intermediate basic reproduction number because higher transmissibility boosts immediate infection but also enhances cross-immunity, reducing future susceptibility.
Significance. If the results hold, this work clarifies how immune history shapes recurrent epidemics of antigenically variable pathogens and explains the non-monotonic relationship between transmissibility and epidemic size. The explicit reduction to a recurrence map under the status-based immunity assumption is a strength, enabling transparent derivation of stability conditions and the reported behaviors directly from the model equations.
minor comments (3)
- Abstract: the claim of 'broad parameter ranges' for stable recurrence would be strengthened by a brief statement of the explored ranges or a reference to the relevant figure or section.
- The recurrence map derivation (likely in the methods or results section) is central; ensure the transition from the full SIR system to the map is shown with all intermediate steps for full reproducibility.
- Figure captions: parameter values used in bifurcation or time-series plots should be listed explicitly to allow readers to reproduce the destabilization at high transmission/low escape.
Simulated Author's Rebuttal
We thank the referee for their positive assessment of our manuscript and for recommending minor revision. The referee's summary accurately reflects the core contributions of the work, including the recurrence map under status-based immunity, the stability of recurrent epidemics, and the non-monotonic dependence of epidemic size on the basic reproduction number. No specific major comments were provided in the report.
Circularity Check
No significant circularity identified
full rationale
The paper constructs a recurrent SIR model that explicitly reduces to a recurrence map for susceptibility under the stated status-based immunity assumption. The central results on stable equal-sized epidemics, destabilization at high transmission/low escape, and non-monotonic epidemic size versus R0 are obtained by direct analysis of that map. No equations or steps in the abstract or skeptic summary reduce a prediction to a fitted input by construction, invoke load-bearing self-citations, or smuggle an ansatz via prior work. The derivation chain is self-contained within the model's assumptions and does not exhibit any of the enumerated circularity patterns.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption status-based population immunity
Reference graph
Works this paper leans on
-
[1]
Dynamics of annual influenza a epidemics with immuno-selection
Andreasen, V., 2003. Dynamics of annual influenza a epidemics with immuno-selection. Journal of mathematical biology 46, 504–536
work page 2003
-
[2]
Shaping the phylogenetic tree of influenza by cross-immunity
Andreasen, V., Sasaki, A., 2006. Shaping the phylogenetic tree of influenza by cross-immunity. Theoretical population biology 70, 164–173
work page 2006
-
[3]
Global migration dynamics underlie evolution and persistence of human influenza a (h3n2)
Bedford, T., Cobey, S., Beerli, P., Pascual, M., 2010. Global migration dynamics underlie evolution and persistence of human influenza a (h3n2). PLoS pathogens 6, e1000918
work page 2010
-
[4]
Biggerstaff, M., Cauchemez, S., Reed, C., Gambhir, M., Finelli, L., 2014. Estimates of the reproduction number for seasonal, pandemic, and zoonotic influenza: a systematic review of the literature. BMC infectious diseases 14, 1–20
work page 2014
-
[5]
Influenza drift and epidemic size: the race between generating and escaping immunity
Boni, M.F., Gog, J.R., Andreasen, V., Christiansen, F.B., 2004. Influenza drift and epidemic size: the race between generating and escaping immunity. Theoretical population biology 65, 179–191
work page 2004
-
[6]
The sirc model and influenza a
Casagrandi, R., Bolzoni, L., Levin, S.A., Andreasen, V., 2006. The sirc model and influenza a. Mathematical biosciences 200, 152–169
work page 2006
-
[7]
Immune history and influenza virus susceptibility
Cobey, S., Hensley, S.E., 2017. Immune history and influenza virus susceptibility. Current opinion in virology 22, 105–111
work page 2017
-
[8]
Complexity of the basic reproduction number (r0)
Delamater, P.L., Street, E.J., Leslie, T.F., Yang, Y.T., Jacobsen, K.H., 2019. Complexity of the basic reproduction number (r0). Emerging infectious diseases 25, 1
work page 2019
-
[9]
Mathematical tools for understanding infectious disease dynamics
Diekmann, O., Heesterbeek, H., Britton, T., 2013. Mathematical tools for understanding infectious disease dynamics. volume 7. Princeton University Press
work page 2013
-
[10]
Antibodylandscapes after influenza virus infection or vaccination
Fonville,J.M.,Wilks,S.,James,S.L.,Fox,A.,Ventresca,M.,Aban,M.,Xue,L.,Jones,T.,Le,N.,Pham,Q.,etal.,2014. Antibodylandscapes after influenza virus infection or vaccination. Science 346, 996–1000
work page 2014
-
[11]
Forecasting epidemiological and evolutionary dynamics of infectious diseases
Gandon, S., Day, T., Metcalf, C.J.E., Grenfell, B.T., 2016. Forecasting epidemiological and evolutionary dynamics of infectious diseases. Trends in ecology & evolution 31, 776–788
work page 2016
-
[12]
Dynamics and selection of many-strain pathogens
Gog, J.R., Grenfell, B.T., 2002. Dynamics and selection of many-strain pathogens. Proceedings of the National Academy of Sciences 99, 17209–17214
work page 2002
-
[13]
Unifying the epidemiological and evolutionary dynamics of pathogens
Grenfell, B.T., Pybus, O.G., Gog, J.R., Wood, J.L., Daly, J.M., Mumford, J.A., Holmes, E.C., 2004. Unifying the epidemiological and evolutionary dynamics of pathogens. Science 303, 327–332
work page 2004
-
[14]
Integrative modeling of seasonal influenza evolution via ai-powered antigenic cartography
Ito, J., Kawakubo, S., Unno, H., Strange, A., Lytras, S., Okumura, K., Lilley, A., Harvey, R., Lewis, N., Sato, K., 2025. Integrative modeling of seasonal influenza evolution via ai-powered antigenic cartography. Biorxiv , 2025–08
work page 2025
-
[15]
Antigenic evolution of dengue viruses over 20 years
Katzelnick,L.C.,CoelloEscoto,A.,Huang,A.T.,Garcia-Carreras,B.,Chowdhury,N.,MaljkovicBerry,I.,Chavez,C.,Buchy,P.,Duong,V., Dussart, P., et al., 2021. Antigenic evolution of dengue viruses over 20 years. Science 374, 999–1004
work page 2021
-
[16]
Modeling infectious diseases in humans and animals
Keeling, M.J., Rohani, P., 2011. Modeling infectious diseases in humans and animals. Princeton university press
work page 2011
-
[17]
Acontributiontothemathematicaltheoryofepidemics
Kermack,W.O.,McKendrick,A.G.,1927. Acontributiontothemathematicaltheoryofepidemics. Proceedingsoftheroyalsocietyoflondon. Series A, Containing papers of a mathematical and physical character 115, 700–721
work page 1927
-
[18]
PLoS computational biology 3, e159
Kryazhimskiy,S.,Dieckmann,U.,Levin,S.A.,Dushoff,J.,2007.Onstate-spacereductioninmulti-strainpathogenmodels,withanapplication to antigenic drift in influenza a. PLoS computational biology 3, e159
work page 2007
-
[19]
Antigenic escape is accelerated by the presence of immunocompromised hosts
Kumata, R., Sasaki, A., 2022. Antigenic escape is accelerated by the presence of immunocompromised hosts. Proceedings of the Royal Society B 289, 20221437
work page 2022
-
[20]
Generality of the final size formula for an epidemic of a newly invading infectious disease
Ma, J., Earn, D.J., 2006. Generality of the final size formula for an epidemic of a newly invading infectious disease. Bulletin of mathematical biology 68, 679–702
work page 2006
-
[21]
Markov,P.V.,Ghafari,M.,Beer,M.,Lythgoe,K.,Simmonds,P.,Stilianakis,N.I.,Katzourakis,A.,2023. Theevolutionofsars-cov-2. Nature Reviews Microbiology 21, 361–379
work page 2023
-
[22]
Populationimmunitypredictsevolutionarytrajectoriesofsars-cov-2
Meijers,M.,Ruchnewitz,D.,Eberhardt,J.,Łuksza,M.,Lässig,M.,2023. Populationimmunitypredictsevolutionarytrajectoriesofsars-cov-2. Cell 186, 5151–5164
work page 2023
-
[23]
Metcalf, C.J.E., Ferrari, M., Graham, A.L., Grenfell, B.T., 2015. Understanding herd immunity. Trends in immunology 36, 753–755
work page 2015
-
[24]
Sars-cov-2 evolution on a dynamic immune landscape
Raharinirina,N.A.,Gubela,N.,Börnigen,D.,Smith,M.R.,Oh,D.Y.,Budt,M.,Mache,C.,Schillings,C.,Fuchs,S.,Dürrwald,R.,etal.,2025. Sars-cov-2 evolution on a dynamic immune landscape. Nature 639, 196–204
work page 2025
-
[25]
Reyné, B., Kamiya, T., Djidjou-Demasse, R., Alizon, S., Sofonea, M.T., 2025. Leaky or polarised immunity: Non-markovian modelling highlights the impact of immune memory assumptions. PLOS Computational Biology 21, e1013399
work page 2025
-
[26]
Asimpleinfluenzamodelwithcomplicateddynamics
Roberts,M.,Hickson,R.I.,McCaw,J.M.,Talarmain,L.,2019. Asimpleinfluenzamodelwithcomplicateddynamics. Journalofmathematical biology 78, 607–624
work page 2019
-
[27]
The global circulation of seasonal influenza a (h3n2) viruses
Russell,C.A.,Jones,T.C.,Barr,I.G.,Cox,N.J.,Garten,R.J.,Gregory,V.,Gust,I.D.,Hampson,A.W.,Hay,A.J.,Hurt,A.C.,etal.,2008. The global circulation of seasonal influenza a (h3n2) viruses. Science 320, 340–346
work page 2008
-
[28]
Immuno-epidemiology and the predictability of viral evolution
Saad-Roy, C.M., Metcalf, C.J.E., Grenfell, B.T., 2022. Immuno-epidemiology and the predictability of viral evolution. Science 376, 1161– 1162
work page 2022
-
[29]
Antigenicescapeselectsfortheevolutionofhigherpathogentransmissionandvirulence
Sasaki,A.,Lion,S.,Boots,M.,2022. Antigenicescapeselectsfortheevolutionofhigherpathogentransmissionandvirulence. Natureecology & evolution 6, 51–62
work page 2022
-
[30]
Smith,D.J.,Lapedes,A.S.,DeJong,J.C.,Bestebroer,T.M.,Rimmelzwaan,G.F.,Osterhaus,A.D.,Fouchier,R.A.,2004.Mappingtheantigenic and genetic evolution of influenza virus. Science 305, 371–376
work page 2004
-
[31]
Synchrony, waves, and spatial hierarchies in the spread of influenza
Viboud, C., Bjørnstad, O.N., Smith, D.L., Simonsen, L., Miller, M.A., Grenfell, B.T., 2006. Synchrony, waves, and spatial hierarchies in the spread of influenza. Science 312, 447–451
work page 2006
-
[32]
Five challenges in modelling interacting strain dynamics
Wikramaratna, P.S., Kucharski, A., Gupta, S., Andreasen, V., McLean, A.R., Gog, J.R., 2015. Five challenges in modelling interacting strain dynamics. Epidemics 10, 31–34. R. Kumata et al.:Preprint submitted to ElsevierPage 11 of 11
work page 2015
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.