A two-patch epidemic model with nonlinear reinfection
Pith reviewed 2026-05-24 18:37 UTC · model grok-4.3
The pith
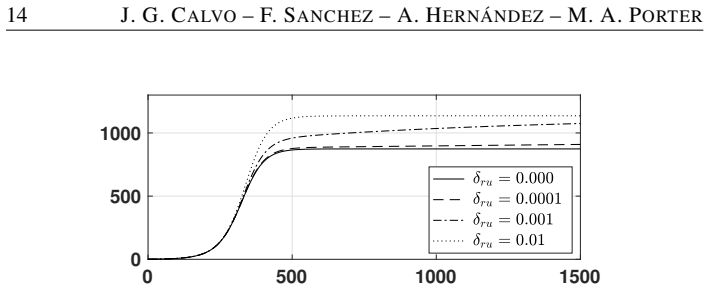
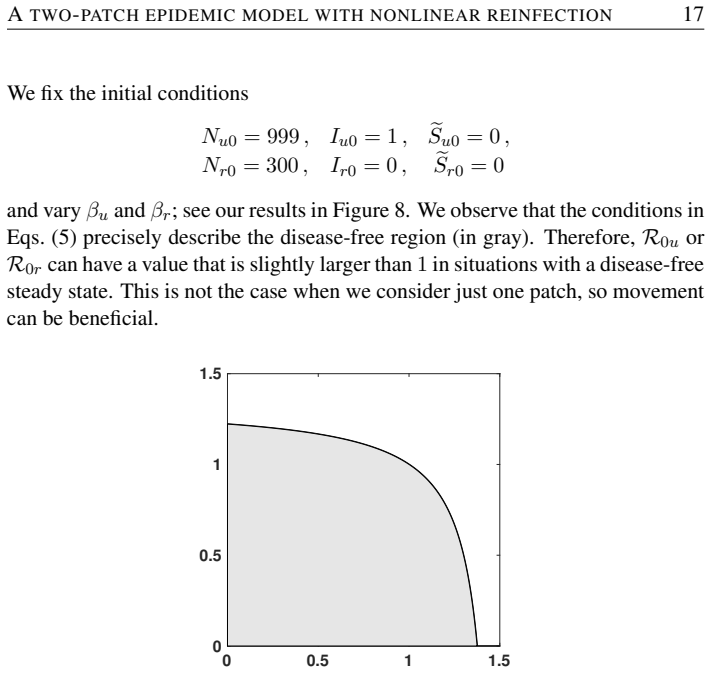
Movement between urban and rural patches can enlarge the stability region of the disease-free equilibrium.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
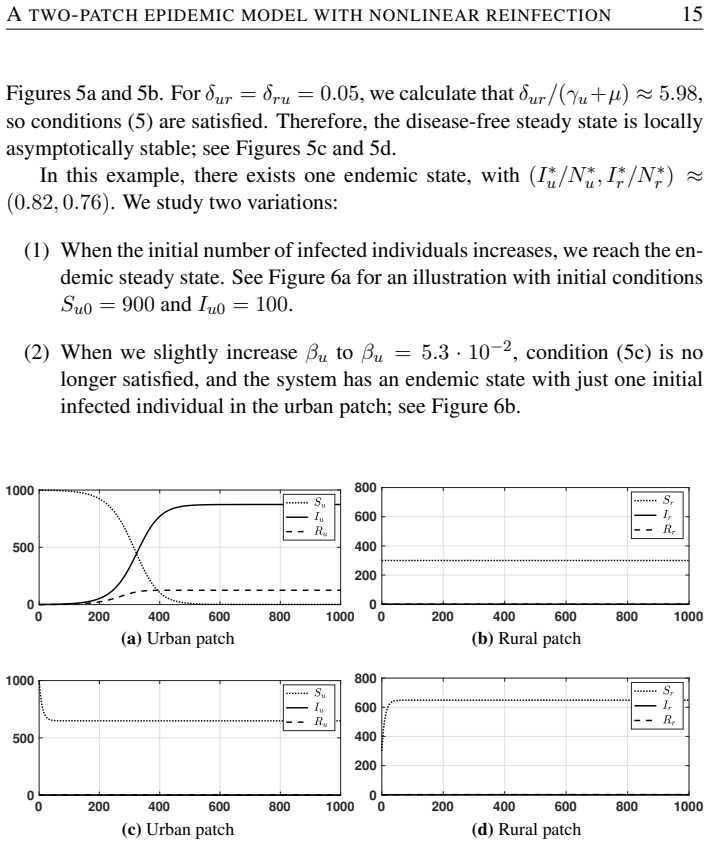
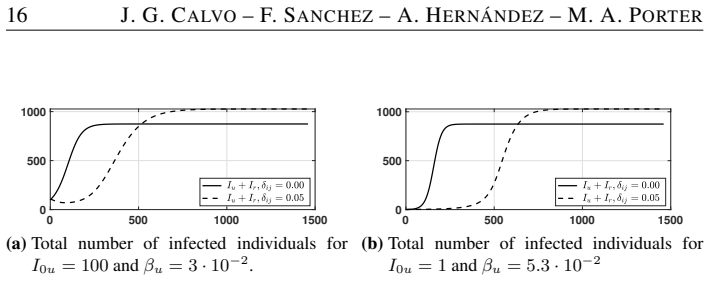
The authors formulate an SI~S compartmental model with nonlinear reinfection in two patches representing urban and rural populations connected by movement. They compute patch-specific basic reproduction numbers, determine conditions for the local stability of the disease-free steady state, and identify when endemic states exist. Their analysis and simulations demonstrate that population movement can enlarge the region of stability of the disease-free steady state.
What carries the argument
The two-patch structure with movement rates between patches, allowing for the calculation of reproduction numbers and stability analysis in the presence of nonlinear reinfection.
Load-bearing premise
The model relies on a specific nonlinear reinfection function and assumes constant movement rates in a minimal two-patch setup adequately represent urban-rural differences.
What would settle it
A simulation or real data set where increasing movement rates between patches decreases rather than increases the stability region of the disease-free state would contradict the claim.
Figures








read the original abstract
The propagation of infectious diseases and its impact on individuals play a major role in disease dynamics, and it is important to incorporate population heterogeneity into efforts to study diseases. As a simplistic but illustrative example, we examine interactions between urban and rural populations in the dynamics of disease spreading. Using a compartmental framework of susceptible--infected--susceptible ($\mathrm{SI\widetilde{S}}$) dynamics with some level of immunity, we formulate a model that allows nonlinear reinfection. We investigate the effects of population movement in the simplest scenario: a case with two patches, which allows us to model movement between urban and rural areas. To study the dynamics of the system, we compute a basic reproduction number for each population (urban and rural). We also compute steady states, determine the local stability of the disease-free steady state, and identify conditions for the existence of endemic steady states. From our analysis and computational experiments, we illustrate that population movement plays an important role in disease dynamics. In some cases, it can be rather beneficial, as it can enlarge the region of stability of a disease-free steady state.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript formulates a two-patch SI~S compartmental model with nonlinear reinfection to study urban-rural epidemic dynamics under constant movement rates. It computes separate basic reproduction numbers for each patch, determines steady states, analyzes local stability of the disease-free equilibrium, derives conditions for endemic equilibria, and uses numerical experiments to conclude that population movement can enlarge the stability region of the disease-free state.
Significance. If the stability claims hold rigorously, the result that movement between patches can beneficially enlarge the disease-free stability region would be of moderate interest for heterogeneous-population epidemic modeling, as it provides a concrete illustration of movement effects beyond standard single-patch thresholds.
major comments (2)
- [Abstract] Abstract: the basic reproduction numbers are stated to be computed separately for each population, yet the local stability of the disease-free equilibrium in a two-patch model with movement is governed by the spectral radius of the next-generation matrix for the full coupled system (four infected compartments linked by movement). Separate per-patch thresholds omit cross-patch transmission and therefore cannot reliably support the central claim that varying movement rates enlarges the stability region.
- [Analysis sections] Analysis of steady states and stability (likely §3–4): without explicit construction of the next-generation matrix for the coupled system or proof that the per-patch R0 values determine the global threshold, the reported conditions for disease-free stability and the numerical observation of enlarged stability cannot be verified as following from the model equations.
minor comments (1)
- The functional form of nonlinear reinfection and the assumption of constant movement rates should be stated more explicitly with parameter ranges used in the computational experiments.
Simulated Author's Rebuttal
We thank the referee for their careful review and constructive comments. The concerns about the next-generation matrix for the coupled system are valid, and we will perform a major revision to incorporate the full analysis.
read point-by-point responses
-
Referee: [Abstract] Abstract: the basic reproduction numbers are stated to be computed separately for each population, yet the local stability of the disease-free equilibrium in a two-patch model with movement is governed by the spectral radius of the next-generation matrix for the full coupled system (four infected compartments linked by movement). Separate per-patch thresholds omit cross-patch transmission and therefore cannot reliably support the central claim that varying movement rates enlarges the stability region.
Authors: We agree that the stability threshold for the coupled two-patch model must be determined by the spectral radius of the next-generation matrix applied to the full four-dimensional infected subsystem that includes movement. The per-patch reproduction numbers were derived under the assumption of isolated patches and do not incorporate cross-patch transmission. In the revision we will rewrite the abstract to state the coupled threshold and note that per-patch quantities are provided only for comparison with the isolated case. revision: yes
-
Referee: [Analysis sections] Analysis of steady states and stability (likely §3–4): without explicit construction of the next-generation matrix for the coupled system or proof that the per-patch R0 values determine the global threshold, the reported conditions for disease-free stability and the numerical observation of enlarged stability cannot be verified as following from the model equations.
Authors: The manuscript indeed presents stability conditions based on the separate per-patch reproduction numbers without constructing the next-generation matrix for the coupled system. Consequently the analytic claims and the numerical observation that movement enlarges the disease-free stability region cannot be rigorously justified from the model equations as written. We will add an explicit next-generation matrix for the four infected compartments, derive the spectral-radius threshold, and re-analyze both the local stability of the disease-free equilibrium and the numerical experiments using the corrected threshold. revision: yes
Circularity Check
No significant circularity; derivation is self-contained
full rationale
The paper formulates an SI~S model on two patches, computes per-patch reproduction numbers, finds steady states, and analyzes local stability of the disease-free equilibrium using standard next-generation methods on the stated equations. No step reduces a claimed result (e.g., movement enlarging the stability region) to a fitted parameter or self-citation by construction. The analysis is independent of the target claims and does not invoke load-bearing self-citations or ansatzes smuggled from prior work by the same authors.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The SĨS compartmental framework with nonlinear reinfection accurately represents the disease process under study.
- domain assumption A two-patch structure with movement rates between patches is sufficient to capture urban-rural heterogeneity.
Reference graph
Works this paper leans on
-
[1]
Costa Rica Once Again Un- der Malaria Alert
Alvarado, L. (2018) “Costa Rica Once Again Un- der Malaria Alert”, in https://news.co.cr/ Rev.Mate.Teor .Aplic.(ISSN print: ????-????; online: ????-????) V ol. ??(?): ?–?, Month Year A TWO -PATCH EPIDEMIC MODEL WITH NONLINEAR REINFECTION 21 costa-rica-once-again-under-malaria-alert/73681 , accessed 25/04/2019
work page 2018
-
[2]
Vector-borne diseases mod- els with residence times — A Lagrangian perspective
Bichara, D.; Castillo-Chavez, C. (2016) “Vector-borne diseases mod- els with residence times — A Lagrangian perspective”, Math. Biosci. 281:128–138
work page 2016
-
[3]
Brauer, R.; Castillo-Chavez, C. (2012) Mathematical Models in Population Biology and Epidemiology , 2nd edition, Springer-Verlag (Providence, RI, USA)
work page 2012
-
[4]
Available from: https://www.cdc.gov/sars/ index.html
Center for Disease Control and Prevention: Severe Acute Respiratory Sys- tem (SARS), 2019. Available from: https://www.cdc.gov/sars/ index.html
work page 2019
-
[5]
Center for Disease Control and Prevention: Ebola (Ebola Virus Disease) ,
-
[6]
Available from: https://www.cdc.gov/vhf/ebola/ history/2014-2016-outbreak/index.html#anchor_ 1515001446180
work page 2014
-
[7]
Available from: https://www.cdc.gov/measles/index.html
Center for Disease Control and Prevention: Measles (Rubeola) , 2019. Available from: https://www.cdc.gov/measles/index.html
work page 2019
-
[8]
Patterns of Movement and Risk of HIV Infection in Rural Zimbabwe
Coffee, M.P.; Garnett, G.P.; Mlilo, M.; V oeten, H.A.C.M.; Chandiwana, S.; Gregson, S. (2005) “Patterns of Movement and Risk of HIV Infection in Rural Zimbabwe”, J. Infect. Dis. 191(1):S159–S167
work page 2005
-
[9]
Chowell G.; Fenimore P.W.; Castillo-Garsow M.A.; Castillo-Chavez C. (2004) “SARS outbreaks in Ontario, Hong Kong and Singapore: The role of diagnosis and isolation as a control mechanism”, J. Theor . Biol. 224(1):1–8
work page 2004
-
[10]
DeV ore, R.A. (1986) “Approximation of functions”, Proc. Sympos. Appl. Math. 36:34–56
work page 1986
-
[11]
Competitive exclusion in a vector–host model for the dengue fever
Feng, Z; Velasco-Hernández, J.X. (1997) “Competitive exclusion in a vector–host model for the dengue fever”, J. Math. Biol. 35:423–544
work page 1997
-
[12]
A model for tubercu- losis with exogenous reinfection
Feng, Z.; Castillo-Chavez, C.; Capurro, A.F. (2000) “A model for tubercu- losis with exogenous reinfection”, Theor . Popul. Biol.57(3):235–247
work page 2000
-
[13]
The Poincaré–Miranda theorem and viability con- dition
Frankowska, H. (2018) “The Poincaré–Miranda theorem and viability con- dition”, J. Math. Anal. Appl. 463:832–837. 22 J. G. C ALVO – F. SANCHEZ – A. H ERNÁNDEZ – M. A. P ORTER
work page 2018
-
[14]
Glynn, J.R.; Murray, J.; Bester, A.; Nelson, G.; Shearer, S; Sonnenberg, P. (2008) “Effects of duration of HIV infection and secondary tuberculosis transmission on tuberculosis incidence in the South African gold mines”, AIDS 22(4):1859–1867
work page 2008
-
[15]
Antibody-Independent Immunity to Reinfection Malaria in B-Cell-Deficient Mice
Grun, J.L.; Weidanz, W.P. (1983) “Antibody-Independent Immunity to Reinfection Malaria in B-Cell-Deficient Mice”, Infect. and Immun. 41(3):1197–1204
work page 1983
-
[16]
Treno, A.J.; Gruenewald, P.; Remer, L.G.; Johnson, F.; LaScala, E.A. (2007) “Examining multi-level relationships between bars, hostility and aggression: social selection and social influence”, Addiction 103:66–77
work page 2007
-
[17]
Kulpa, W. (1997) “The Poincaré–Miranda theorem”, Amer . Math. Month. 104(6):545–550
work page 1997
-
[18]
The role of residence times in two- patch dengue transmission dynamics and optimal strategies
Lee, S.; Castillo-Chavez, C. (2015) “The role of residence times in two- patch dengue transmission dynamics and optimal strategies”,J. Theor . Biol. 374:152–164
work page 2015
-
[19]
Manore, C.A.; Hickmann, K.S.; Xu, S.; Wearing, H.J.; Hyman, J.M. (2014) “Comparing dengue and chikungunya emergence and endemic transmis- sion in A. aegypti and A. albopictus”, J. Theor . Biol.356:174–191
work page 2014
-
[20]
Malaria on the move: Human population movement and malaria transmission
Martens, P.; Hall, L. (2000) “Malaria on the move: Human population movement and malaria transmission”, Emerg. Infect. Diseas. 6(2):103– 109
work page 2000
-
[21]
Vertical transmission in a two-strain model of dengue fever
Murillo, D.; Holechek, S.; Murillo, A.; Sanchez, F.; Castillo-Chavez, C. (2014) “Vertical transmission in a two-strain model of dengue fever”, Lett. Biomath. 1(2):249–271
work page 2014
-
[22]
(1996) Medical Microbiology, 4th edition
Crutcher, J.M.; Hoffman, S.L. (1996) Medical Microbiology, 4th edition. https://www.ncbi.nlm.nih.gov/books/NBK8584/
work page 1996
-
[23]
Epidemic processes in complex networks
Pastor-Satorras, R.; Castellano, C.; Van Mieghem, P.; Vespignani, A. (2015) “Epidemic processes in complex networks”, Rev. Mod. Phys. 87(3), 925–979
work page 2015
-
[24]
The role of vaccination in the control of SARS
Gjorgjieva, J; Smith, K; Chowell, G.; Sanchez, F; Snyder, J; Castillo- Chavez, C. (2005) “The role of vaccination in the control of SARS”, Math. Biosci. Eng. 2(4):753–769. Rev.Mate.Teor .Aplic.(ISSN print: ????-????; online: ????-????) V ol. ??(?): ?–?, Month Year A TWO -PATCH EPIDEMIC MODEL WITH NONLINEAR REINFECTION 23
work page 2005
-
[25]
Drinking as an epidemic — A simple mathematical model with recovery and relapse
Sanchez, F.; Wang, X.; Castillo-Chavez, C.; Gorman, D.; Gruenewald, P. (2007) “Drinking as an epidemic — A simple mathematical model with recovery and relapse”, In: Witkiewitz, K.A., Marlatt, G.A., eds. Therapist’s Guide to Evidence-Based Relapse Prevention Academic Press (Cambridge, MA, USA), 353–368
work page 2007
-
[26]
Change in host be- havior and its impact on the transmission dynamics of dengue
Sanchez, F.; Murillo, D.; Castillo-Chavez, C. (2012) “Change in host be- havior and its impact on the transmission dynamics of dengue”, in Interna- tional Symposium on Mathematical and Computational Biology , (Ed. R.P. Mondaini), BIOMAT 2011:191–203
work page 2012
-
[27]
Sanchez, F.; Calvo, J.G.; Segura, E.; Feng, Z. (2018) “A partial differ- ential equation model with age-structure and nonlinear recidivism: Condi- tions for a backward bifurcation and a general numerical implementation”, arXiv:1712.09105
-
[28]
Raves, clubs and ecstasy: The impact of peer pressure
Song, B.; Castillo-Garsow, M.; Rios-Soto, K.; Mejran, M.; Henso, L.; Castillo-Chavez, C. (2006) “Raves, clubs and ecstasy: The impact of peer pressure”, Math. Biosci. Eng. 3(1):249–266
work page 2006
-
[29]
Different types of backward bifurcations due to density-dependent treatments
Song, B.; Du, W.; Lou, J. (2013) “Different types of backward bifurcations due to density-dependent treatments”, Math. Biosci. Eng. 10(5–6):1651– 1668
work page 2013
-
[30]
Models for Dengue transmission and control
Sanchez, F.; Engman, M.; Harrington, L; Castillo-Chávez, C. (2006) “Models for Dengue transmission and control”, Mathematical Studies on Human Disease Dynamics, 311–326, Contemp. Math., 410, Amer. Math. Soc., Providence, RI, USA
work page 2006
-
[31]
On a generalization of the Miranda Theorem and its application to boundary value problems
Szyma ´nska-De ¸bowska, K. (2015) “On a generalization of the Miranda Theorem and its application to boundary value problems”, J. Diff. Equ. 258(8):2686–2700
work page 2015
-
[32]
The Bolzano–Poicaré–Miranda theorem — Discrete version
Turzánski, M. (2012) “The Bolzano–Poicaré–Miranda theorem — Discrete version”, Topol. Appl. 159:3130–3135
work page 2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.