Effective hyperuniformity in time-integrated stochastic Turing patterns
Pith reviewed 2026-06-26 06:15 UTC · model grok-4.3
The pith
Temporal integration of stochastic configurations in the Levin-Segel model produces effective hyperuniformity with variance scaling as 1/R over expanding ranges near the Turing instability.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
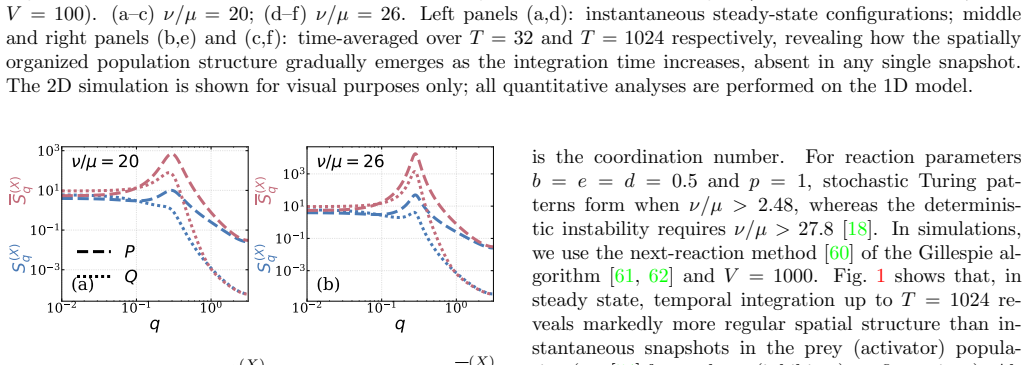
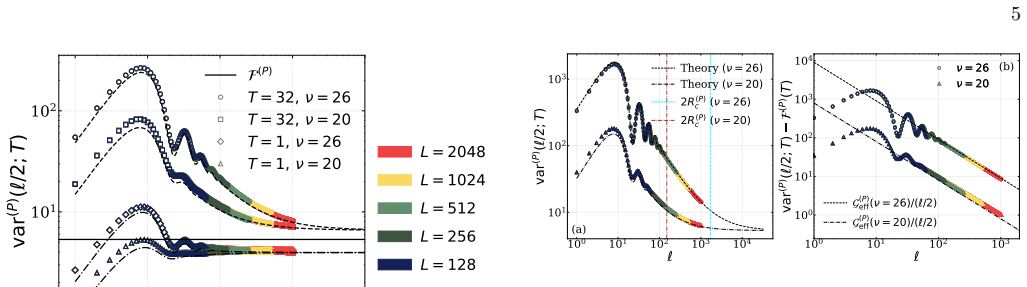
In the Levin-Segel model, temporal integration of stochastic configurations reveals that the intensive number variance in a window of size R ≫ 1 approaches a finite reaction-kinetic floor as 1/R. This scaling persists over a spatial range that grows by orders of magnitude near the Turing instability, yielding an effectively hyperuniform, fine-tuning-free regime previously unidentified in non-conserved multispecies stochastic systems.
What carries the argument
The 1/R approach of intensive number variance to a reaction-kinetic floor under temporal integration of stochastic configurations near the Turing instability.
If this is right
- The hyperuniform scaling appears only after time integration rather than in single snapshots.
- The organized regime extends over dramatically larger distances as the Turing instability is approached.
- The effect occurs in non-conserved multispecies systems without requiring parameter fine-tuning.
- The finite floor is set by reaction kinetics rather than by diffusion or conservation constraints.
Where Pith is reading between the lines
- The same integration approach could be tested in other reaction-diffusion models to check whether the 1/R scaling is generic.
- Experimental time-lapse data from chemical or ecological systems might be reanalyzed to search for similar variance floors.
- Varying the integration window length could map how the effective hyperuniform range depends on observation time.
Load-bearing premise
That temporal integration of the stochastic configurations directly produces the reported variance scaling independently of specific choices for integration timescale, noise strength, or other model details.
What would settle it
Numerical measurements in the Levin-Segel model showing that the intensive number variance fails to approach a finite floor as 1/R or that the scaling range does not expand near the Turing instability.
Figures

read the original abstract
Demographic noise generates stochastic Turing patterns even when reaction-diffusion systems are deterministically stable. We show analytically and verify numerically in the Levin-Segel model that temporal integration of configurations reveals emergent large-scale organization. The intensive number variance in a window of size $R \gg 1$ approaches a finite reaction-kinetic floor as $1/R$, over a spatial range growing by orders of magnitude near the Turing instability. This yields an effectively hyperuniform, fine-tuning-free regime previously unidentified in non-conserved multispecies stochastic systems.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that demographic noise in deterministically stable reaction-diffusion systems generates stochastic Turing patterns, and that temporal integration of these configurations in the Levin-Segel model produces emergent large-scale organization. Analytically and numerically, the intensive number variance in a window of size R ≫ 1 is shown to approach a finite reaction-kinetic floor as 1/R, with the spatial range of this scaling expanding by orders of magnitude near the Turing instability. This is presented as yielding an effectively hyperuniform, fine-tuning-free regime in non-conserved multispecies stochastic systems.
Significance. If the central claim holds, the result is significant for identifying a previously unrecognized hyperuniform regime in non-conserved stochastic reaction-diffusion systems that does not require fine-tuning or conservation. The analytical derivation combined with numerical verification across parameters is a strength. The stress-test concern on dependence of the 1/R scaling on integration timescale T and noise strength does not land on the manuscript, as the paper explicitly derives the floor from the reaction kinetics and verifies the scaling numerically for appropriate T relative to reaction/diffusion timescales and across noise amplitudes.
minor comments (1)
- The notation for the integration window T and its relation to the reaction and diffusion timescales could be clarified in §3 to make the parameter regime explicit for readers.
Simulated Author's Rebuttal
We thank the referee for their positive assessment of the manuscript, accurate summary of the central claims, and recommendation for minor revision. The referee correctly notes the significance of the effectively hyperuniform regime in non-conserved stochastic reaction-diffusion systems and that the analytical derivation of the reaction-kinetic floor, together with the numerical verification, addresses potential concerns about dependence on integration timescale and noise strength.
Circularity Check
No circularity: derivation from model dynamics is self-contained
full rationale
The abstract and provided excerpts frame the 1/R variance scaling as an emergent consequence of temporal integration applied to the stochastic Levin-Segel reaction-diffusion equations near the Turing point. No equations or claims reduce a 'prediction' to a fitted parameter by construction, invoke self-citations as load-bearing uniqueness theorems, or smuggle ansatzes via prior work. The result is presented as derived from the underlying stochastic dynamics without redefinition or statistical forcing. This matches the default expectation of non-circularity for papers whose central claim rests on explicit model analysis rather than reparameterization.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Demographic noise generates stochastic Turing patterns even in deterministically stable reaction-diffusion systems
Reference graph
Works this paper leans on
-
[1]
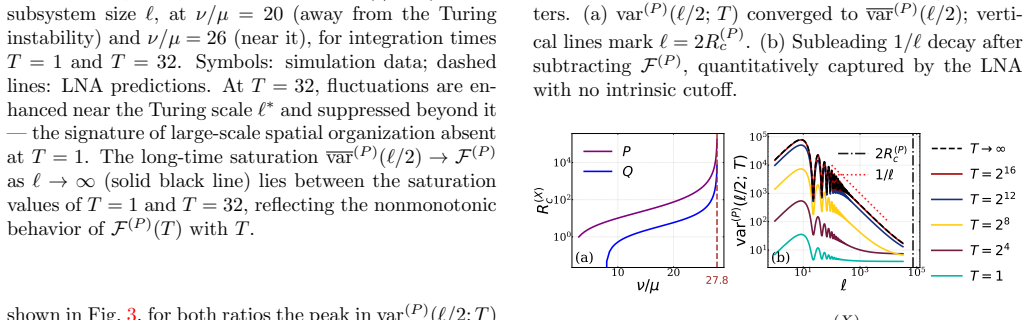
Symbols: simula- tion data forL= 128–2048 (color codes as in Fig
andν/µ= 26 (R (P) c ≈850). Symbols: simula- tion data forL= 128–2048 (color codes as in Fig. 3); dashed lines: LNA predictions with no fitting parame- ters. (a) var (P) (ℓ/2;T) converged to var(P) (ℓ/2); verti- cal lines markℓ= 2R (P) c . (b) Subleading 1/ℓdecay after subtractingF (P) , quantitatively captured by the LNA with no intrinsic cutoff. 10 20 / ...
2048
-
[2]
J. D. Murray,Mathematical Biology II: Spatial Models and Biomedical Applications, 3rd ed., Interdisciplinary Applied Mathematics, Vol. 18 (Springer, New York, 2003)
2003
-
[3]
M. C. Cross and P. C. Hohenberg, Rev. Mod. Phys.65, 851 (1993)
1993
-
[4]
Camazine, J.-L
S. Camazine, J.-L. Deneubourg, N. R. Franks, J. Sneyd, G. Theraulaz, and E. Bonabeau,Self-Organization in Bi- ological Systems(Princeton University Press, Princeton, NJ, USA, 2003)
2003
-
[5]
Evasion of tipping in complex systems through spatial pattern formation
M. Rietkerk, R. Bastiaansen, S. Banerjee, J. van de Koppel, M. Baudena, and A. Doelman, Science374, 10.1126/science.abj0359 (2021)
-
[6]
Turing, Philos
A. Turing, Philos. Trans. R. Soc. London, Ser. B237, 37 (1952)
1952
-
[7]
Kondo and T
S. Kondo and T. Miura, Science329, 1616 (2010)
2010
-
[8]
Rietkerk and J
M. Rietkerk and J. van de Koppel, Trends Ecol. Evol. 23, 169 (2008)
2008
-
[9]
Weber,Philosophy of Developmental Biology(Cam- bridge University Press, Cambridge, England, UK, 2022)
M. Weber,Philosophy of Developmental Biology(Cam- bridge University Press, Cambridge, England, UK, 2022)
2022
-
[10]
Smith, Nature381, 367 (1996)
J. Smith, Nature381, 367 (1996)
1996
-
[11]
J. B. Gurdon and P.-Y. Bourillot, Nature413, 797 (2001)
2001
-
[12]
Wolpert, Trends Genet.12, 359 (1996)
L. Wolpert, Trends Genet.12, 359 (1996)
1996
-
[13]
L. S. Tsimring, Rep. Prog. Phys.77, 026601 (2014)
2014
-
[14]
Lande, S
R. Lande, S. Engen, and B.-E. Saether,Stochastic Pop- ulation Dynamics in Ecology and Conservation(Oxford University Press, 2003)
2003
-
[15]
A. J. McKane and T. J. Newman, Phys. Rev. E70, 041902 (2004)
2004
-
[16]
C. A. Lugo and A. J. McKane, Phys. Rev. E78, 051911 (2008)
2008
-
[17]
A. J. McKane and T. J. Newman, Phys. Rev. Lett.94, 218102 (2005)
2005
-
[18]
Butler and N
T. Butler and N. Goldenfeld, Phys. Rev. E80, 030902 (2009)
2009
-
[19]
Butler and N
T. Butler and N. Goldenfeld, Phys. Rev. E84, 011112 (2011)
2011
-
[20]
Biancalani, D
T. Biancalani, D. Fanelli, and F. Di Patti, Phys. Rev. E 81, 046215 (2010)
2010
-
[21]
Asslani, F
M. Asslani, F. Di Patti, and D. Fanelli, Phys. Rev. E86, 046105 (2012)
2012
-
[22]
Karig, K
D. Karig, K. M. Martini, T. Lu, N. A. DeLateur, N. Gold- enfeld, and R. Weiss, Proc. Natl. Acad. Sci. U.S.A.115, 6572 (2018)
2018
-
[23]
Di Patti, L
F. Di Patti, L. Lavacchi, R. Arbel-Goren, L. Schein- Lubomirsky, D. Fanelli, and J. Stavans, PLoS Biol.16, e2004877 (2018)
2018
-
[24]
Di Patti, Y
F. Di Patti, Y. Ugartechea Chirino, R. Arbel-Goren, T. Sharon, A. Castillo, E. Alvarez-Buylla, D. Fanelli, and J. Stavans, Proc. Natl. Acad. Sci. U.S.A.120, e2309616120 (2023)
2023
-
[25]
Di Patti, R
F. Di Patti, R. Arbel-Goren, D. Fanelli, and J. Stavans, Phys. Rep.1179, 1 (2026)
2026
-
[26]
Wolpert, J
L. Wolpert, J. Theor. Biol.25, 1 (1969)
1969
-
[27]
Pag` es and S
F. Pag` es and S. Kerridge, Trends Genet.16, 40 (2000)
2000
-
[28]
H. L. Ashe and J. Briscoe, Development133, 385 (2006)
2006
-
[29]
F. Borgogno, P. D’Odorico, F. Laio, and L. Ridolfi, Rev. Geophys.47, 10.1029/2007RG000256 (2009)
-
[30]
B. D. Harfe, P. J. Scherz, S. Nissim, H. Tian, A. P. McMahon, and C. J. Tabin, Cell118, 517 (2004)
2004
-
[31]
Dessaud, L
E. Dessaud, L. L. Yang, K. Hill, B. Cox, F. Ulloa, A. Ribeiro, A. Mynett, B. G. Novitch, and J. Briscoe, Nature450, 717 (2007), 18046410
2007
-
[32]
Dessaud, A
E. Dessaud, A. P. McMahon, and J. Briscoe, Develop- 7 ment135, 2489 (2008)
2008
-
[33]
Sagner and J
A. Sagner and J. Briscoe, WIREs Dev. Biol.6, e271 (2017)
2017
-
[34]
K. Ogle, J. J. Barber, G. A. Barron-Gafford, L. P. Bent- ley, J. M. Young, T. E. Huxman, M. E. Loik, and D. T. Tissue, Ecol. Lett.18, 221 (2015)
2015
-
[35]
J. F. Johnstone, C. D. Allen, J. F. Franklin, L. E. Frelich, B. J. Harvey, P. E. Higuera, M. C. Mack, R. K. Meente- meyer, M. R. Metz, G. L. W. Perry, T. Schoennagel, and M. G. Turner, Front. Ecol. Environ.14, 369 (2016)
2016
-
[36]
Torquato and F
S. Torquato and F. H. Stillinger, Phys. Rev. E68, 041113 (2003)
2003
-
[37]
Torquato, Phys
S. Torquato, Phys. Rep.745, 1 (2018)
2018
-
[38]
Y. Jiao, T. Lau, H. Hatzikirou, M. Meyer-Hermann, J. C. Corbo, and S. Torquato, Phys. Rev. E89, 022721 (2014)
2014
-
[39]
Y. Liu, D. Chen, J. Tian, W. Xu, and Y. Jiao, Phys. Rev. Lett.133, 028401 (2024)
2024
-
[40]
Ge, Proc
Z. Ge, Proc. Natl. Acad. Sci. U.S.A.120, e2306514120 (2023)
2023
-
[41]
Huang, W
M. Huang, W. Hu, S. Yang, Q.-X. Liu, and H. P. Zhang, Proc. Natl. Acad. Sci. U.S.A.118, e2100493118 (2021)
2021
-
[42]
Mayer, V
A. Mayer, V. Balasubramanian, T. Mora, and A. M. Wal- czak, Proc. Natl. Acad. Sci. U.S.A.112, 5950 (2015)
2015
-
[43]
Tunable Hyperuniformity in Cellular Structures (2024), [Online; accessed 3. Jun. 2026]
2024
-
[44]
Siegert, L
S. Siegert, L. Kanari, and M. C. U¸ car, Self-organized tiling generates tissue-scale hyperuniformity during de- velopment (2026), 2026.04.30.721955
2026
-
[45]
Ma and S
Z. Ma and S. Torquato, J. Appl. Phys.121, 244904 (2017)
2017
-
[46]
Zheng, M
Y. Zheng, M. A. Klatt, and H. L¨ owen, Phys. Rev. Res. 6, L032056 (2024)
2024
-
[47]
Ballestero, A
E. Ballestero, A. Duclos, A. Barbacci, and V. Romero- Garc´ ıa, Phys. Rev. E112, L062401 (2025)
2025
-
[48]
Hexner and D
D. Hexner and D. Levine, Phys. Rev. Lett.114, 110602 (2015)
2015
-
[49]
Mukherjee and P
A. Mukherjee and P. Pradhan, Phys. Rev. E107, 024109 (2023)
2023
-
[50]
Hexner and D
D. Hexner and D. Levine, Phys. Rev. Lett.118, 020601 (2017)
2017
-
[51]
Mukherjee, D
A. Mukherjee, D. Tapader, A. Hazra, and P. Pradhan, Phys. Rev. E110, 024119 (2024)
2024
-
[52]
Maire and L
R. Maire and L. Chaix, J. Chem. Phys.163, 214507 (2025)
2025
-
[53]
T. Agranov, N. Wiegenfeld, O. Karin, and B. D. Si- mons, Self-organized hyperuniformity in a minimal model of population dynamics (2025), arXiv:2509.08077 [cond- mat.stat-mech]
arXiv 2025
-
[54]
N. G. Van Kampen,Stochastic Processes in Physics and Chemistry(North Holland, 2007)
2007
-
[55]
See Supplemental Material at [URL will be inserted by publisher] for the full analytical derivations and addi- tional numerical results
-
[56]
T. G. Kurtz, J. Appl. Probab.7, 49 (1970)
1970
-
[57]
T. G. Kurtz, J. Appl. Probab.8, 344 (1971)
1971
-
[58]
Cross and H
M. Cross and H. Greenside,Pattern Formation and Dy- namics in Nonequilibrium Systems(Cambridge Univer- sity Press, Cambridge, England, UK, 2009)
2009
-
[59]
Biancalani, F
T. Biancalani, F. Jafarpour, and N. Goldenfeld, Phys. Rev. Lett.118, 018101 (2017)
2017
-
[60]
S. A. Levin and L. A. Segel, Nature259, 659 (1976)
1976
-
[61]
M. A. Gibson and J. Bruck, J. Phys. Chem. A104, 1876 (2000)
2000
-
[62]
D. T. Gillespie, Annu. Rev. Phys. Chem. , 35 (2007)
2007
-
[63]
D. T. Gillespie, A. Hellander, and L. R. Petzold, J. Chem. Phys.138, 10.1063/1.4801941 (2013)
-
[64]
HereR≡ℓ/2 denotes the hyperspherical sampling- window radius in general dimension, reducing in our 1D simulations to half the subsystem lengthℓ
-
[65]
Biancalani, T
T. Biancalani, T. Galla, and A. J. McKane, Phys. Rev. E84, 026201 (2011)
2011
-
[66]
A. F. Peralta and R. Toral, Chaos28, 106303 (2018)
2018
-
[67]
DLMF,NIST Digital Library of Mathematical Functions, https://dlmf.nist.gov/, Release 1.2.7 of 2026-06-15, f. W. J. Olver, A. B. Olde Daalhuis, D. W. Lozier, B. I. Schneider, R. F. Boisvert, C. W. Clark, B. R. Miller, B. V. Saunders, H. S. Cohl, and M. A. McClain, eds. 8 Supplemental Material: Effective hyperuniformity in time-integrated stochastic Turing ...
2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.