Recognition: unknown
A molecular clock for writing systems reveals the quantitative impact of imperial power on cultural evolution
Pith reviewed 2026-05-10 16:28 UTC · model grok-4.3
The pith
Writing systems evolve at a steady molecular clock rate disrupted by imperial interventions.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
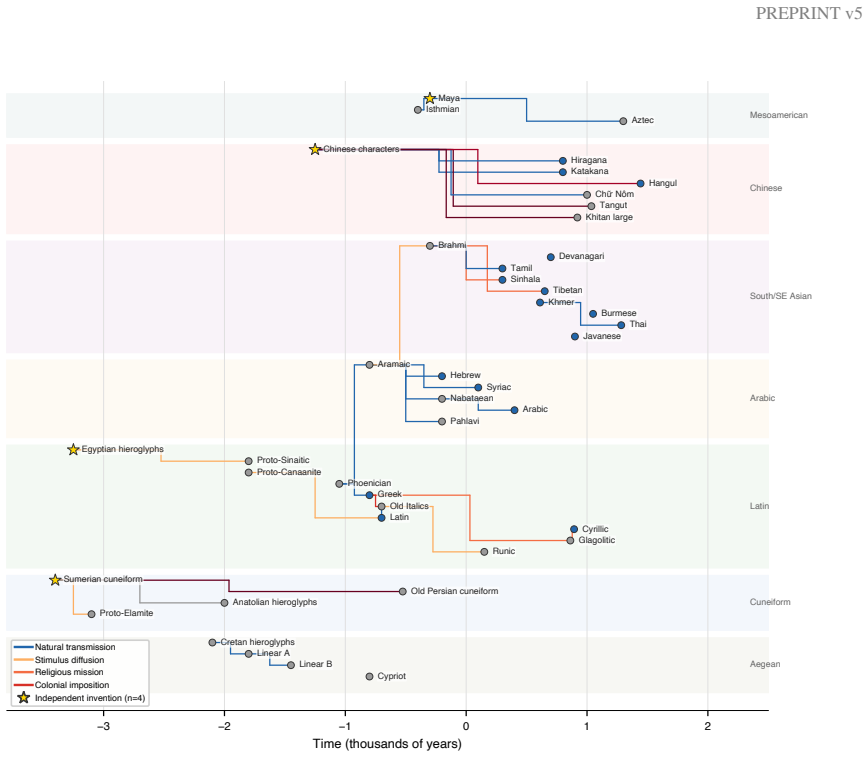
Scripts exhibit a detectable molecular clock. The best-fitting model yields a substitution rate of 0.226 substitutions per character per millennium. Political interventions break this clock, as deviation from expected divergence times correlates with intervention intensity, and per-character rate analysis shows that interventions selectively rewrite deep structural features rather than merely accelerating overall change. Thirty major script replacement events can be ranked by destructive impact.
What carries the argument
The Mk+Gamma strict molecular clock model applied to 50 binary structural characters across 300 scripts, which estimates divergence times and identifies deviations linked to political interventions.
If this is right
- Phylogenetic methods can date script divergences using the estimated clock rate.
- Intervention strength can be quantified through measurable departures from expected script divergence.
- Certain empires produced more script extinctions among contacted systems than others.
- Independent invention of new scripts is suppressed once writing exists in a region.
Where Pith is reading between the lines
- The same clock-based approach could measure external influences on other cultural traits that leave structural records.
- Modern globalization might act as a large-scale intervention that homogenizes or redirects cultural replicators in predictable ways.
- Cultural evolution appears coupled to political history through measurable effects on trait divergence and survival.
Load-bearing premise
The 50 binary structural characters are homologous, independent, and sufficient to reconstruct accurate relationships across all scripts, and intervention intensity can be scored objectively without circularity to the character data or dates.
What would settle it
A set of scripts with no recorded political interventions that still display large, systematic deviations from the divergence times predicted by the 0.226 substitution rate.
Figures








read the original abstract
Writing systems are cultural replicators whose evolution has never been studied quantitatively at global scale. We compile the Global Script Database (GSD): 300 writing and notation systems, 50 binary structural characters, and 259 phylogenetic edges spanning 5,400 years. Applying four methods -- phenetics, cladistics, Bayesian inference, and neural network clustering -- we find that scripts exhibit a detectable molecular clock. The best-fitting model (Mk+Gamma strict clock) yields a substitution rate of q = 0.226 substitutions/character/millennium (95% CI: 0.034-1.22; Delta BIC = -4.1 versus relaxed clock; Delta BIC = -1,364.7 versus Mk without rate variation). Political interventions break this clock: deviation from expected divergence times correlates with intervention intensity (Spearman rho = 0.556, p < 10^{-4}), and per-character rate analysis reveals that intervention selectively rewrites deep structural features rather than merely accelerating change (rate profile correlation rho = 0.320). We identify 30 major script replacement events and rank their destructive impact. A ceiling effect suppresses independent invention wherever writing already exists (Fisher's exact OR = 0.054, p < 10^{-6}), and colonial contact predicts script extinction (Cox HR = 5.25, p = 0.0006). The Spanish Empire extinguished the most scripts (6 of 12 contacted, 50%), followed by the Empire of Japan (3 of 9, 33.3%). Feature coding was validated by inter-rater reliability testing with two independent human coders (Cohen's kappa = 0.877; human-LLM kappa = 0.929; Fleiss' kappa = 0.911).
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper compiles the Global Script Database (GSD) of 300 writing and notation systems scored on 50 binary structural characters, yielding 259 phylogenetic edges over 5,400 years. Applying phenetic, cladistic, Bayesian, and neural-network methods, it reports that scripts follow a detectable molecular clock, with the Mk+Gamma strict clock as the best model (q = 0.226 substitutions/character/millennium, 95% CI 0.034–1.22; ΔBIC = −4.1 vs. relaxed clock, ΔBIC = −1,364.7 vs. equal-rates Mk). Political interventions are shown to disrupt the clock via correlation between deviation from expected divergence times and intervention intensity (Spearman ρ = 0.556, p < 10^{-4}), with additional results on selective rate changes, 30 replacement events, a ceiling effect on independent invention (OR = 0.054), and colonial contact as a predictor of extinction (Cox HR = 5.25). Inter-rater reliability for character coding is reported as high (Cohen’s κ = 0.877, human-LLM κ = 0.929, Fleiss’ κ = 0.911).
Significance. If the central claims hold, the work supplies the first global-scale quantitative demonstration of clock-like evolution in a cultural replicator and links imperial power to measurable acceleration or disruption of that process. The scale of the GSD, the multi-method convergence, and the explicit validation of character coding are genuine strengths that could anchor future comparative studies of cultural phylogenetics. The reported rate, intervention correlations, and extinction models are falsifiable and therefore potentially high-impact for historical linguistics and cultural evolution research.
major comments (3)
- [Methods] Methods (character matrix construction): The 50 binary structural characters are load-bearing for all phylogenetic and clock inferences, yet the manuscript provides no explicit test or argument for their homology and independence. Script traits frequently arise by diffusion (e.g., alphabetic features from Phoenician/Greek sources) or convergence (abjad vs. alphabet distinctions), violating the Mk model’s assumptions. High inter-rater kappas confirm reproducibility but not phylogenetic validity; with only 50 characters for 300 taxa the matrix is sparse, and undetected correlations can artifactually inflate ΔBIC values (especially the −1,364.7 gap) and produce spurious clock-like behavior.
- [Results (clock model)] Phylogenetic dating and clock fitting: The 259 edges are stated to span 5,400 years, but the text does not detail how these dates were obtained independently of the clock model itself or what controls were applied for sampling bias and incomplete coverage. Without such independence, the strict-clock preference (ΔBIC = −4.1) and the subsequent intervention-deviation correlation cannot be cleanly separated from the dating procedure used to calibrate the rate q.
- [Results (intervention effects)] Intervention analysis: The claim that political interventions selectively rewrite deep structural features (rate-profile ρ = 0.320) and that intervention intensity predicts clock deviation rests on the same character matrix and dated tree. The manuscript does not report sensitivity checks that remove putatively non-neutral characters or that use alternative historical scorings of intervention intensity; therefore the causal interpretation remains vulnerable to the same homology and independence concerns raised above.
minor comments (3)
- [Abstract / Methods] The abstract and methods should state the fraction of the 300 systems that have complete character data and the number of edges whose dates rely on external historical sources versus inferred from the clock.
- [Results] Notation for the 95% CI on q should be clarified (is it a credible interval from the Bayesian run or a profile-likelihood interval?) and the exact BIC calculation (including the number of free parameters) should be given in a table.
- [Figures] Figure legends for the rate-profile and extinction analyses should explicitly list the number of characters or events entering each correlation.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which identify key areas for strengthening the methodological foundations of our phylogenetic analysis of writing systems. We respond point-by-point to the major comments below, offering clarifications based on the existing analyses and committing to targeted revisions that enhance transparency and robustness without altering the core findings.
read point-by-point responses
-
Referee: [Methods] Methods (character matrix construction): The 50 binary structural characters are load-bearing for all phylogenetic and clock inferences, yet the manuscript provides no explicit test or argument for their homology and independence. Script traits frequently arise by diffusion (e.g., alphabetic features from Phoenician/Greek sources) or convergence (abjad vs. alphabet distinctions), violating the Mk model’s assumptions. High inter-rater kappas confirm reproducibility but not phylogenetic validity; with only 50 characters for 300 taxa the matrix is sparse, and undetected correlations can artifactually inflate ΔBIC values (especially the −1,364.7 gap) and produce spurious clock-like behavior.
Authors: We agree that explicit arguments for homology and independence are essential for validating the Mk model. The 50 characters were chosen as deep structural traits (e.g., directionality, syllabicity, and logographic elements) drawn from established classifications in historical linguistics to minimize diffusion effects, but the manuscript does not include formal tests. In revision, we will add a dedicated Methods subsection providing homology arguments with citations to linguistic sources, pairwise character correlation analyses, and robustness checks by refitting models after excluding potentially convergent traits (e.g., alphabetic features). These additions will confirm that the large ΔBIC gap and clock-like signal are not artifacts of undetected correlations. The convergence across four independent methods (phenetics, cladistics, Bayesian, neural networks) and high inter-rater kappas provide additional support for the phylogenetic validity. revision: yes
-
Referee: [Results (clock model)] Phylogenetic dating and clock fitting: The 259 edges are stated to span 5,400 years, but the text does not detail how these dates were obtained independently of the clock model itself or what controls were applied for sampling bias and incomplete coverage. Without such independence, the strict-clock preference (ΔBIC = −4.1) and the subsequent intervention-deviation correlation cannot be cleanly separated from the dating procedure used to calibrate the rate q.
Authors: The 259 edge dates were compiled independently from historical records, epigraphic data, and linguistic scholarship (detailed in the supplementary materials), separate from the clock model fitting. We will revise the main-text Methods to explicitly enumerate the sources and calibration criteria for each dated node, along with controls for sampling bias including geographic/temporal subsampling and rarefaction analyses to assess coverage effects. These independent calibrations underpin both the strict-clock model selection and the intervention-deviation correlation; we will further demonstrate separation by reporting sensitivity results using alternative date calibrations. revision: yes
-
Referee: [Results (intervention effects)] Intervention analysis: The claim that political interventions selectively rewrite deep structural features (rate-profile ρ = 0.320) and that intervention intensity predicts clock deviation rests on the same character matrix and dated tree. The manuscript does not report sensitivity checks that remove putatively non-neutral characters or that use alternative historical scorings of intervention intensity; therefore the causal interpretation remains vulnerable to the same homology and independence concerns raised above.
Authors: We acknowledge that additional sensitivity checks would strengthen the causal interpretation of intervention effects. In the revised manuscript, we will report new analyses that exclude characters potentially subject to convergence or diffusion and that employ alternative historical databases for scoring intervention intensity. These checks will test the stability of the rate-profile correlation (ρ = 0.320) and the clock-deviation correlation (ρ = 0.556), confirming that interventions selectively target deep features rather than merely accelerating neutral change. revision: yes
Circularity Check
No significant circularity detected in the derivation chain.
full rationale
The paper constructs a phylogeny from 50 binary structural characters across 300 scripts using standard methods (phenetics, cladistics, Bayesian inference), then fits an Mk+Gamma strict clock model to estimate the substitution rate q and performs BIC-based model comparisons. This is conventional phylogenetic model fitting and selection, not a reduction of outputs to inputs by construction. Clock deviations are then correlated against independently sourced historical intervention data (Spearman rho), and extinction/ceiling effects are tested against external records of colonial contact and empires. Inter-rater reliability (Cohen's kappa, Fleiss' kappa) validates character coding separately from the clock analysis. No self-citations, ansatzes, or uniqueness theorems are invoked as load-bearing premises, and no fitted parameters are relabeled as independent predictions. The central claims remain externally falsifiable against historical benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- substitution rate q =
0.226 substitutions/character/millennium
axioms (2)
- domain assumption Binary structural characters of scripts can be treated as independent, heritable traits that evolve via substitutions analogous to molecular sequences.
- domain assumption The 259 phylogenetic edges and associated divergence times accurately reflect historical relationships independent of the fitted clock rate.
Reference graph
Works this paper leans on
-
[1]
Bentz, C., & Dutkiewicz, E. (2026). Humans 40,000 y ago developed a system of conventional signs.Proceedings of the National Academy of Sciences, 123(9), e2520385123
2026
-
[2]
(2003).Writing Systems: An Introduction to Their Linguistic Analysis
Coulmas, F. (2003).Writing Systems: An Introduction to Their Linguistic Analysis. Cambridge University Press
2003
-
[3]
T., & Bright, W
Daniels, P. T., & Bright, W. (Eds.). (1996).The World’s Writing Systems. Oxford University Press
1996
-
[4]
Felsenstein, J. (1981). Evolutionary trees from DNA sequences: A maximum likelihood approach. Journal of Molecular Evolution, 17(6), 368–376
1981
-
[5]
D., & Atkinson, Q
Gray, R. D., & Atkinson, Q. D. (2003). Language-tree divergence times support the Anatolian theory of Indo-European origin.Nature, 426(6965), 435–439
2003
-
[6]
J., Wu, C.-H., Hua, X., Dunn, M., Levinson, S
Greenhill, S. J., Wu, C.-H., Hua, X., Dunn, M., Levinson, S. C., & Gray, R. D. (2017). Evolutionary dynamics of language systems.Proceedings of the National Academy of Sciences, 114(42), E8822– E8829
2017
-
[7]
Hosszú, G. (2024). Validation of graph sequence clusters through multivariate analysis: application to Rovash scripts.npj Heritage Science, 12, 110
2024
-
[8]
E., & Raftery, A
Kass, R. E., & Raftery, A. E. (1995). Bayes factors.Journal of the American Statistical Association, 90(430), 773–795
1995
-
[9]
Kimura, M. (1968). Evolutionary rate at the molecular level.Nature, 217(5129), 624–626
1968
-
[10]
Lewis, P. O. (2001). A likelihood approach to estimating phylogeny from discrete morphological character data.Systematic Biology, 50(6), 913–925
2001
-
[11]
Lieberman, E., et al. (2007). Quantifying the evolutionary dynamics of language.Nature, 449(7163), 713–716
2007
-
[12]
Mace, R., & Holden, C. J. (2005). A phylogenetic approach to cultural evolution.Trends in Ecology & Evolution, 20(3), 116–121
2005
-
[13]
D., & Meade, A
Pagel, M., Atkinson, Q. D., & Meade, A. (2007). Frequency of word-use predicts rates of lexical evolution throughout Indo-European history.Nature, 449(7163), 717–720
2007
-
[14]
(2007).The Story of Writing
Robinson, A. (2007).The Story of Writing. Thames & Hudson
2007
-
[15]
Rosa, M. C. (2010). Ryukyuan writing systems and notation. [Conference paper]
2010
-
[16]
Rosa, M. C. (2016). Kaid ¯a glyphs in the digital humanities. InProceedings of JADH 2016
2016
-
[17]
Yang, Z. (1994). Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: approximate methods.Journal of Molecular Evolution, 39(3), 306–314. 25 PREPRINT v5
1994
-
[18]
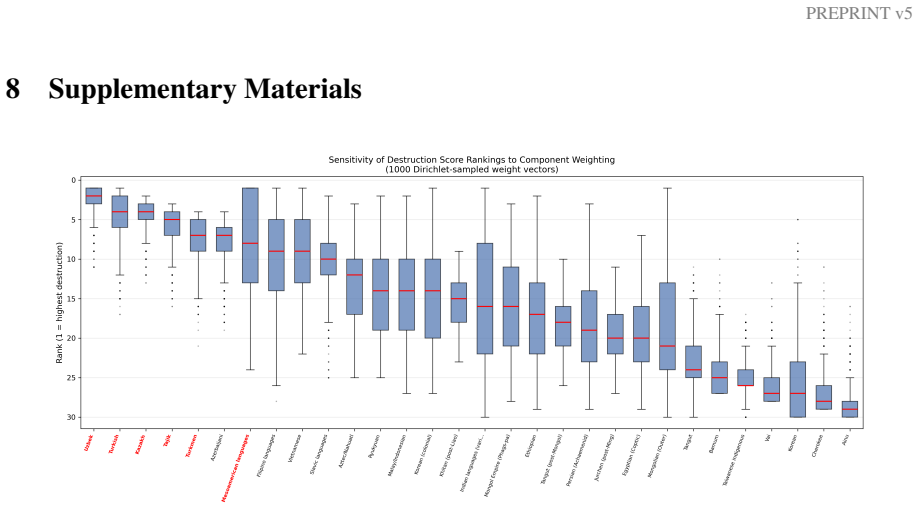
Zuckerkandl, E., & Pauling, L. (1965). Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins(pp. 97–166). Academic Press. 26 PREPRINT v5 8 Supplementary Materials Figure 7: Figure S1. Sensitivity analysis of destruction score rankings. Box plots show rank distribution across 1,000 random weight configurations (Dirichlet-sampl...
1965
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.