Recognition: unknown
Local growth laws determine global shape of molluscan shells
Pith reviewed 2026-05-08 12:41 UTC · model grok-4.3
The pith
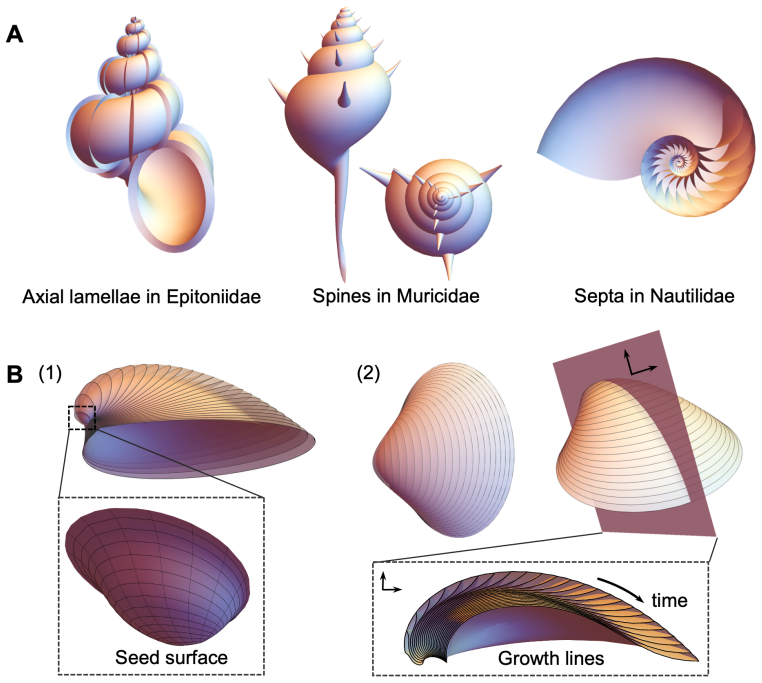
Molluscan shell shapes are produced by repeated application of a local growth law at the edge, reducing nearly all varieties to a scaling factor, an orientation vector, and the protoconch edge curve.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
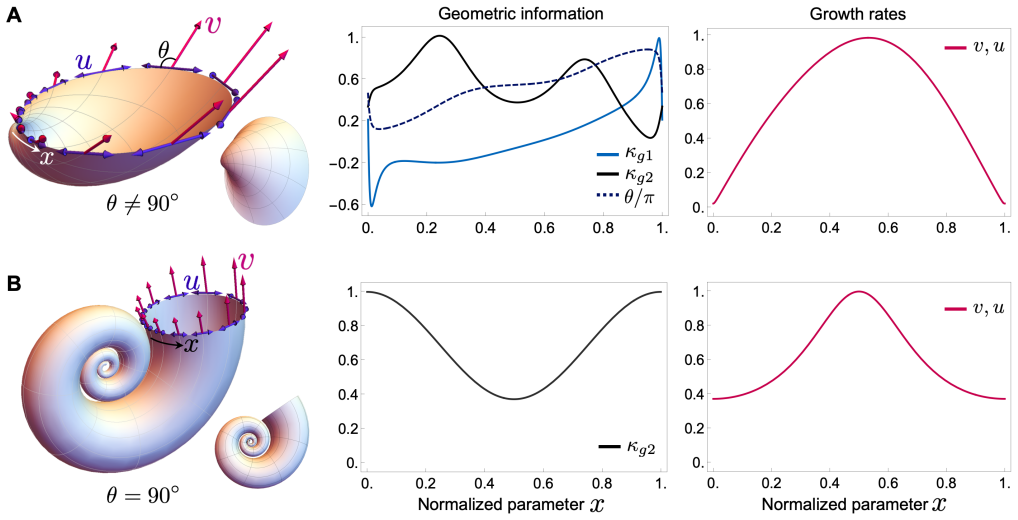
The shape of the shell is generated by the action of a Lie group on a protoconch, where the group admits a representation determined solely by local geometry at the growing edge. This representation shows that the shapes of nearly all known molluscan shells are described by three parameters: a scalar scaling, a vector for orientation, and the curve of the protoconch edge.
What carries the argument
The Lie group action on the protoconch, represented so that its generators depend only on local edge geometry.
If this is right
- Shell morphology becomes independent of external conditions once the growth law and initial protoconch are fixed.
- The three parameters provide a low-dimensional coordinate system in which phylogenetic relationships among shell forms can be read directly.
- The same local-to-global construction supplies a generative method for designing other complex curved surfaces without global coordination.
- Variations in the protoconch edge curve alone account for most observed diversity while keeping the growth rule unchanged.
Where Pith is reading between the lines
- The same three-parameter description could be tested against quantitative measurements of growth rates along the mantle edge in living mollusks.
- If the local rule is encoded genetically, mutations affecting only the edge curve would produce large shape changes while preserving the growth law itself.
- The construction may extend to other accretive biological structures whose final form is built by successive addition at a moving boundary.
- Parametric models derived from this representation could be used to generate families of shell-like objects for structural engineering or materials design.
Load-bearing premise
Growth follows one fixed rule that depends only on local geometry at the current edge and is applied repeatedly without any biological machinery that monitors or corrects the overall shape.
What would settle it
A single well-documented mollusk shell whose three-dimensional form cannot be recovered by any choice of scaling, orientation vector, and protoconch edge curve generated from repeated local growth steps.
Figures


read the original abstract
Molluscan shells come in various shapes and sizes. Despite this diversity, each species produces a shell with a characteristic shape that is independent of environmental conditions. We seek to understand this robust complexity. We are guided by two principles in the spirit of D'Arcy Thompson. First, the growth is governed by the repeated and continuous application of a fixed growth law, even as the shell evolves in overall shape, without any complex biological machinery to monitor and control the growth. Second, the growth law depends solely on local geometry at the shell's growing edge. The first principle naturally leads to the mathematical statement that the shape of the shell is generated by the action of a Lie group on a protoconch. The second naturally leads to a particular representation of the Lie group. We use this representation to show that the shapes of nearly all known molluscan shells can be described by essentially three parameters: a scalar (scaling), a vector (orientation), and a curve (edge of the protoconch). We relate these parameters to the phylogenetic tree. In addition to the morphogenetic insight, our results potentially point to a new approach to engineering complex structures.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that molluscan shell shapes arise from the repeated application of a fixed local growth law depending only on geometry at the aperture edge. This is formalized mathematically as the action of a specific Lie group on an initial protoconch curve. The authors conclude that this framework shows nearly all known shell shapes are captured by three parameters (a scalar scaling factor, an orientation vector, and the protoconch edge curve), relate the parameters to phylogeny, and suggest applications to engineering complex structures.
Significance. If the reduction from arbitrary local rules to a three-parameter Lie-group action is rigorously established, the work would supply a parsimonious, parameter-efficient model linking local geometry to global morphology. This aligns with D'Arcy Thompson-style principles and could constrain evolutionary explanations while offering a template for designing self-similar curved objects.
major comments (3)
- [Abstract] Abstract: the assertion that the representation 'shows' nearly all known shells are described by three parameters supplies no dataset, quantitative fitting criterion, or verification procedure. Without these, the empirical scope of the central claim cannot be assessed.
- [§3] §3 (derivation of the Lie-group representation): the map from a general local growth law (normal velocity depending on local curvature, torsion, or position) to a Lie group whose only continuous parameters are scaling and orientation is asserted but not derived. No explicit generators or proof excluding additional invariants or non-commuting elements is given, so it remains unclear whether the parameter count is generally restricted to three.
- [§5] §5 (phylogenetic comparison): the reported correlation between the extracted parameters and the phylogenetic tree lacks details on how parameters were measured from real shells, the statistical method used, or controls for confounding variables such as shell size.
minor comments (2)
- [Abstract] The phrase 'essentially three parameters' is imprecise because the protoconch edge is a curve (infinite-dimensional); clarify whether it is treated as a fixed template or as additional free parameters.
- [§2] Notation for the Lie-group action and the orientation vector should be defined explicitly at first use rather than introduced informally.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed comments on our manuscript. We address each major comment point by point below, providing the strongest honest defense of the work while acknowledging where revisions are needed to improve clarity and rigor. We have revised the manuscript accordingly.
read point-by-point responses
-
Referee: [Abstract] Abstract: the assertion that the representation 'shows' nearly all known shells are described by three parameters supplies no dataset, quantitative fitting criterion, or verification procedure. Without these, the empirical scope of the central claim cannot be assessed.
Authors: The claim in the abstract is a mathematical one arising from the structure of the Lie group action: local geometry-dependent growth laws generate transformations that reduce shell shapes to parameterization by a scalar, an orientation vector, and the initial protoconch curve. This is a theoretical reduction shown via the group properties rather than an empirical fit to data. We have revised the abstract to explicitly state the theoretical nature of the demonstration. To provide verification, we have added illustrative examples in a new figure showing how the three parameters generate representative shell forms from the literature. revision: partial
-
Referee: [§3] §3 (derivation of the Lie-group representation): the map from a general local growth law (normal velocity depending on local curvature, torsion, or position) to a Lie group whose only continuous parameters are scaling and orientation is asserted but not derived. No explicit generators or proof excluding additional invariants or non-commuting elements is given, so it remains unclear whether the parameter count is generally restricted to three.
Authors: We acknowledge that §3 presented the reduction concisely without full detail. The local growth law, defined as a normal velocity depending only on local geometry, induces infinitesimal generators that form a Lie algebra closed under commutation; due to the locality and lack of explicit position or global dependence, the algebra is spanned solely by the scaling generator and the three rotation generators (corresponding to the orientation vector), with no additional independent invariants. We have expanded §3 with the explicit generators and a proof sketch confirming the three-parameter restriction holds for the class of local laws considered. revision: yes
-
Referee: [§5] §5 (phylogenetic comparison): the reported correlation between the extracted parameters and the phylogenetic tree lacks details on how parameters were measured from real shells, the statistical method used, or controls for confounding variables such as shell size.
Authors: We agree that the original presentation lacked sufficient methodological detail. In the revised §5 we have added a full description of parameter extraction (edge detection on shell images followed by optimization to fit scaling and orientation to the aperture curve), the statistical procedure (Mantel test correlating parameter distances with phylogenetic distances), and controls (normalization by shell size with size included as a covariate). We also discuss potential confounders such as environmental effects and measurement error. revision: yes
Circularity Check
No significant circularity; derivation is self-contained from stated principles.
full rationale
The paper begins from two explicit principles (repeated fixed growth law implying Lie group action on protoconch; local-geometry dependence implying a particular group representation) and derives the three-parameter description as a mathematical consequence. No step reduces by construction to a fitted input, self-citation, or renamed empirical pattern; the classification of shells follows from the representation rather than presupposing it. The chain is independent of the target result and does not rely on load-bearing self-citations or ansatzes imported from prior work by the same authors.
Axiom & Free-Parameter Ledger
free parameters (2)
- scaling scalar
- orientation vector
axioms (2)
- domain assumption Growth is governed by the repeated and continuous application of a fixed growth law even as the shell evolves in overall shape, without any complex biological machinery to monitor and control the growth.
- domain assumption The growth law depends solely on local geometry at the shell's growing edge.
Reference graph
Works this paper leans on
-
[1]
Interdependence of cell growth and gene expression: origins and consequences.Science, 330(6007):1099–1102, 2010
Matthew Scott, Carl W Gunderson, Eduard M Mateescu, Zhongge Zhang, and Terence Hwa. Interdependence of cell growth and gene expression: origins and consequences.Science, 330(6007):1099–1102, 2010
2010
-
[2]
Growth rate-dependent global effects on gene expression in bacteria.Cell, 139(7):1366–1375, 2009
Stefan Klumpp, Zhongge Zhang, and Terence Hwa. Growth rate-dependent global effects on gene expression in bacteria.Cell, 139(7):1366–1375, 2009
2009
-
[3]
A scalable pipeline for designing reconfigurable organisms.Proceedings of the National Academy of Sciences, 117(4):1853–1859, 2020
Sam Kriegman, Douglas Blackiston, Michael Levin, and Josh Bongard. A scalable pipeline for designing reconfigurable organisms.Proceedings of the National Academy of Sciences, 117(4):1853–1859, 2020
2020
-
[4]
Growth and morphogenesis of the gastropod shell.Proceedings of the national academy of sciences, 116(14):6878–6883, 2019
Adam B Johnson, Nina S Fogel, and J David Lambert. Growth and morphogenesis of the gastropod shell.Proceedings of the national academy of sciences, 116(14):6878–6883, 2019
2019
-
[5]
The neural origins of shell struc- ture and pattern in aquatic mollusks.Proceedings of the National Academy of Sciences, 106(16):6837–6842, 2009
Alistair Boettiger, Bard Ermentrout, and George Oster. The neural origins of shell struc- ture and pattern in aquatic mollusks.Proceedings of the National Academy of Sciences, 106(16):6837–6842, 2009
2009
-
[6]
Dy- namic morphoskeletons in development.Proceedings of the National Academy of Sciences, 117(21):11444–11449, 2020
Mattia Serra, Sebastian Streichan, Manli Chuai, Cornelis J Weijer, and L Mahadevan. Dy- namic morphoskeletons in development.Proceedings of the National Academy of Sciences, 117(21):11444–11449, 2020
2020
-
[7]
Biomineral- ization in mollusks and brachiopods.https://bioengineering.hyperbook.mcgill.ca/ biomineralization-in-mollusks-and-brachiopods/
Elliott Cole, Gabriel Linaje-Ferrel, Mikaela Phung, and Salma Oraichi. Biomineral- ization in mollusks and brachiopods.https://bioengineering.hyperbook.mcgill.ca/ biomineralization-in-mollusks-and-brachiopods/. Accessed: 2025-08-05
2025
-
[8]
Molluscan shell proteins: primary structure, origin, and evolution.Current topics in developmental biology, 80:209–276, 2007
Frédéric Marin, Gilles Luquet, Benjamin Marie, and Davorin Medakovic. Molluscan shell proteins: primary structure, origin, and evolution.Current topics in developmental biology, 80:209–276, 2007
2007
-
[9]
Mollusc shellomes: past, present and future.Journal of Structural Biology, 212(1):107583, 2020
Frédéric Marin. Mollusc shellomes: past, present and future.Journal of Structural Biology, 212(1):107583, 2020
2020
-
[10]
The physical basis of mollusk shell chiral coiling.Proceedings of the National Academy of Sciences, 118(48):e2109210118, 2021
Régis Chirat, Alain Goriely, and Derek E Moulton. The physical basis of mollusk shell chiral coiling.Proceedings of the National Academy of Sciences, 118(48):e2109210118, 2021
2021
-
[11]
Non-classical crystallization in soft and organic materials.Nature Reviews Materials, 9(4):229–248, 2024
Jingshan S Du, Yuna Bae, and James J De Yoreo. Non-classical crystallization in soft and organic materials.Nature Reviews Materials, 9(4):229–248, 2024
2024
-
[12]
Cambridge University Press, 1917
D’Arcy Wentworth Thompson.On growth and form. Cambridge University Press, 1917
1917
-
[13]
Ontheformationoftheskeletonsofanimalsandotherhardstructuresformedin connexion with living tissues.The British and Foreign Medico-Chirurgical Review, 20(40):451, 1857
GeorgeRainey. Ontheformationoftheskeletonsofanimalsandotherhardstructuresformedin connexion with living tissues.The British and Foreign Medico-Chirurgical Review, 20(40):451, 1857. 11
-
[14]
Number 1
Pieter Harting.Recherches de morphologie synthétique sur la production artificielle de quelques formations calcaires organiques. Number 1. Van der post, 1872
-
[15]
Crystal growth kinetics as an architectural constraint on the evolution of molluscan shells.Proceedings of the National Academy of Sciences, 116(41):20388–20397, 2019
Vanessa Schoeppler, Robert Lemanis, Elke Reich, Tamás Pusztai, László Gránásy, and Igor Zlotnikov. Crystal growth kinetics as an architectural constraint on the evolution of molluscan shells.Proceedings of the National Academy of Sciences, 116(41):20388–20397, 2019
2019
-
[16]
Classical view on nonclassical crystal growth in a biological setting.Journal of the American Chemical Society, 147(1):1–9, 2024
Richard Johannes Best, Deborah Stier, Lucas Kuhrts, and Igor Zlotnikov. Classical view on nonclassical crystal growth in a biological setting.Journal of the American Chemical Society, 147(1):1–9, 2024
2024
-
[17]
Phase-field modeling of biomineralization in mollusks and corals: Microstructure vs formation mechanism.JACS Au, 1(7):1014–1033, 2021
László Gránásy, László Rátkai, Gyula I Tóth, Pupa UPA Gilbert, Igor Zlotnikov, and Tamás Pusztai. Phase-field modeling of biomineralization in mollusks and corals: Microstructure vs formation mechanism.JACS Au, 1(7):1014–1033, 2021
2021
-
[18]
Neuropeptides regulate shell growth in the mediterranean mussel (mytilus galloprovincialis).International Journal of Biological Macromolecules, 281:136500, 2024
Zhi Li, Maoxiao Peng, Rute C Félix, Joao CR Cardoso, and Deborah M Power. Neuropeptides regulate shell growth in the mediterranean mussel (mytilus galloprovincialis).International Journal of Biological Macromolecules, 281:136500, 2024
2024
-
[19]
Growth uncoupling and the relationship between shell growth and metabolism in the soft shell clam mya arenaria.Marine Ecology Progress Series, 158:177–189, 1997
Daniel E Lewis and Robert M Cerrato. Growth uncoupling and the relationship between shell growth and metabolism in the soft shell clam mya arenaria.Marine Ecology Progress Series, 158:177–189, 1997
1997
-
[20]
Geometric analysis of shell coiling: general problems.Journal of paleontology, pages 1178–1190, 1966
David M Raup. Geometric analysis of shell coiling: general problems.Journal of paleontology, pages 1178–1190, 1966
1966
-
[21]
Kinematics of surface growth.Journal of mathematical biology, 35(8):869–907, 1997
R Skalak, DA Farrow, and Anne Hoger. Kinematics of surface growth.Journal of mathematical biology, 35(8):869–907, 1997
1997
-
[22]
Surface growth kinematics via local curve evolution
Derek E Moulton and Alain Goriely. Surface growth kinematics via local curve evolution. Journal of mathematical biology, 68(1):81–108, 2014
2014
-
[23]
Aca- demic press, 1979
Sigurdur Helgason.Differential geometry, Lie groups, and symmetric spaces, volume 80. Aca- demic press, 1979
1979
-
[24]
Dorsoventral decoupling of hox gene expression underpins the diversification of molluscs.Proceedings of the national academy of sciences, 117(1):503–512, 2020
Pin Huan, Qian Wang, Sujian Tan, and Baozhong Liu. Dorsoventral decoupling of hox gene expression underpins the diversification of molluscs.Proceedings of the national academy of sciences, 117(1):503–512, 2020
2020
-
[25]
Dynamic expression of ancient and novel molluscan shell genes during ecological transitions.BMC evolutionary biology, 7(1):160, 2007
Daniel J Jackson, Gert Wörheide, and Bernard M Degnan. Dynamic expression of ancient and novel molluscan shell genes during ecological transitions.BMC evolutionary biology, 7(1):160, 2007
2007
-
[26]
A genome-based phylogeny for mollusca is concordant with fossils and morphology.Science, 387(6737):1001–1007, 2025
Zeyuan Chen, J Antonio Baeza, Chong Chen, Maria Teresa Gonzalez, Vanessa Liz González, Carola Greve, Kevin M Kocot, Pedro Martinez Arbizu, Juan Moles, Tilman Schell, et al. A genome-based phylogeny for mollusca is concordant with fossils and morphology.Science, 387(6737):1001–1007, 2025. 12
2025
-
[27]
Age and growth rate estimations of the commercially fished gastropod buccinum undatum.ICES Journal of Marine Science, 75(6):2129–2144, 2018
Philip R Hollyman, Simon RN Chenery, Melanie J Leng, Vladimir V Laptikhovsky, Charlotte N Colvin, and Christopher A Richardson. Age and growth rate estimations of the commercially fished gastropod buccinum undatum.ICES Journal of Marine Science, 75(6):2129–2144, 2018
2018
-
[28]
Richard L Shelmerdine, Jennifer Adamson, Chevonne H Laurenson, and Beth Leslie. Size vari- ation of the common whelk, buccinum undatum, over large and small spatial scales: potential implications for micro-management within the fishery.Fisheries Research, 86(2-3):201–206, 2007
2007
-
[29]
J. S. Langer. Instabilities and pattern formation in crystal growth.Reviews of Modern Physics, 52:1–28, 1980
1980
-
[30]
The morphological evolution of dendritic microstructures during coarsening.Acta materialia, 54(6):1549–1558, 2006
D Kammer and PW Voorhees. The morphological evolution of dendritic microstructures during coarsening.Acta materialia, 54(6):1549–1558, 2006
2006
-
[31]
Solidification microstructures and solid-state parallels: Recent developments, future directions.Acta Materialia, 57(4):941–971, 2009
Mark Asta, Christoph Beckermann, Alain Karma, Wilfried Kurz, R Napolitano, Mathis Plapp, G Purdy, Michel Rappaz, and Rohit Trivedi. Solidification microstructures and solid-state parallels: Recent developments, future directions.Acta Materialia, 57(4):941–971, 2009
2009
-
[32]
Vegetation enhances curvature-driven dynamics in meandering rivers.Nature Communications, 15(1):1968, 2024
Alvise Finotello, Alessandro Ielpi, Mathieu GA Lapôtre, Eli D Lazarus, Massimiliano Ghinassi, Luca Carniello, Serena Favaro, Davide Tognin, and Andrea D’Alpaos. Vegetation enhances curvature-driven dynamics in meandering rivers.Nature Communications, 15(1):1968, 2024
1968
-
[33]
How multiscale cur- vature couples forces to cellular functions.Nature Reviews Physics, 6(4):246–268, 2024
Marine Luciano, Caterina Tomba, Aurélien Roux, and Sylvain Gabriele. How multiscale cur- vature couples forces to cellular functions.Nature Reviews Physics, 6(4):246–268, 2024
2024
-
[34]
Curvature and rho activation differentially control the alignment of cells and stress fibers.Science advances, 3(9):e1700150, 2017
Nathan D Bade, Randall D Kamien, Richard K Assoian, and Kathleen J Stebe. Curvature and rho activation differentially control the alignment of cells and stress fibers.Science advances, 3(9):e1700150, 2017
2017
-
[35]
3d curvature-instructed endothelial flow response and tissue vascularization.Science advances, 6(38):eabb3629, 2020
Christian Mandrycky, Brandon Hadland, and Ying Zheng. 3d curvature-instructed endothelial flow response and tissue vascularization.Science advances, 6(38):eabb3629, 2020
2020
-
[36]
Geometric control of tissue morphogenesis.Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1793(5):903–910, 2009
Celeste M Nelson. Geometric control of tissue morphogenesis.Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1793(5):903–910, 2009
2009
-
[37]
Marking time with bivalve shells: oxygen isotopes and season of annual increment formation.Palaios, pages 340–346, 1996
Douglas S Jones and Irvy R Quitmyer. Marking time with bivalve shells: oxygen isotopes and season of annual increment formation.Palaios, pages 340–346, 1996
1996
-
[38]
Douglas S Jones. Sclerochronology: reading the record of the molluscan shell: annual growth increments in the shells of bivalve molluscs record marine climatic changes and reveal surprising longevity.American Scientist, 71(4):384–391, 1983
1983
-
[39]
Comprehensive scaling laws across animals, microorganisms and plants.Proceedings of the Royal Society A, 481(2320):20250515, 2025
Huan Liu, Shashank Priya, and Richard D James. Comprehensive scaling laws across animals, microorganisms and plants.Proceedings of the Royal Society A, 481(2320):20250515, 2025
2025
-
[40]
Morphogenic growth 3d printing.Advanced Materials, 37(20):2406265, 2025
Yun Seong Kim, Minjiang Zhu, Mohammad Tanver Hossain, Derrick Sanders, Rohan Shah, Yuan Gao, Jeffrey S Moore, Nancy R Sottos, Randy H Ewoldt, Philippe H Geubelle, et al. Morphogenic growth 3d printing.Advanced Materials, 37(20):2406265, 2025. 13
2025
-
[41]
American Mathematical Soc., 2015
Wolfgang Kühnel.Differential geometry, volume 77. American Mathematical Soc., 2015
2015
-
[42]
University of Toronto Press, 1968
Erwin Kreyszig.Introduction to differential geometry and Riemannian geometry. University of Toronto Press, 1968. 14 Appendix A Geometric preliminaries We recall some basic properties of manifolds (see, for example, [41, 42]). Consider aC3-smooth surface ( two-dimensional manifold embedded in three-dimensional Euclidean space) parametrized by parametersα 1...
1968
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.