The Sensation Modulating Network:Haltability as the architectural ground for object-directed phenomenology
Pith reviewed 2026-06-29 14:56 UTC · model grok-4.3
The pith
Haltability in opponent body dynamics supplies the architecture for object-directed phenomenology without added modules.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
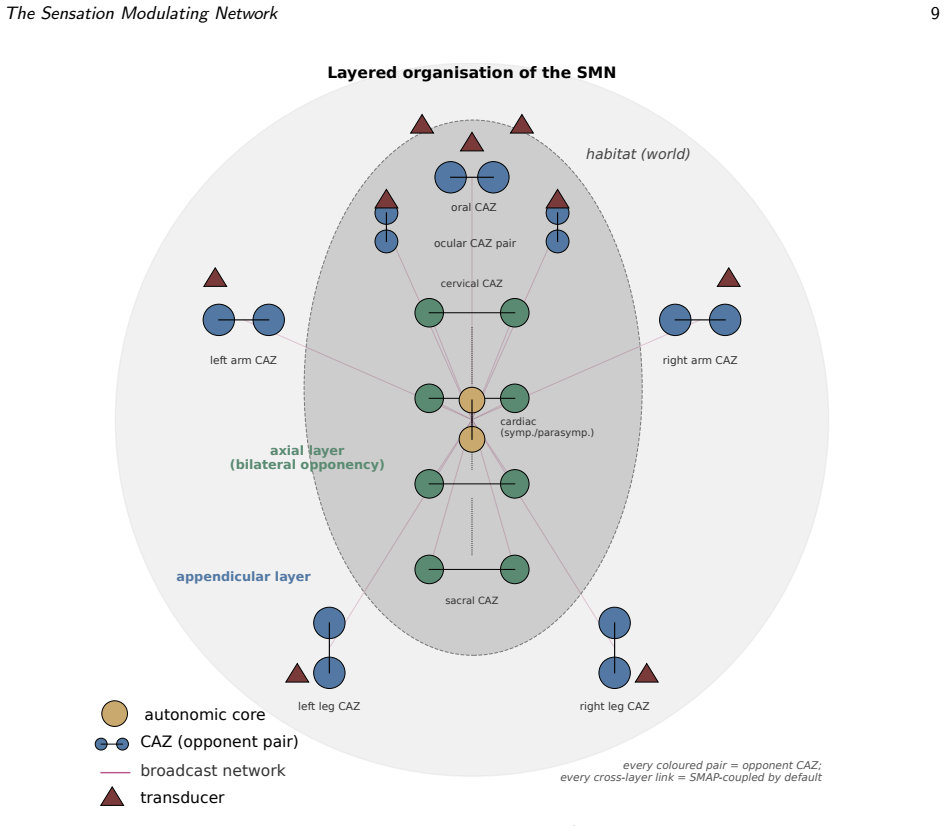
Haltability—the recruitment of antagonistic affordance into co-activated equilibrium—provides the architectural locus that object-directed phenomenology requires: opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness, with no module added on top. The SMN realizes this through sensation modulators and coordinated action zones under a broadcast network, while the dual-signal property of self-modulatable action patterns and the four-level action-pattern hierarchy (Basal, Haltable, Negotiable, Transactional) locate recursion in modifiable dynamics and embodiment in the opponent substrate.
What carries the argument
Haltability, the recruitment of antagonistic affordance into co-activated equilibrium within the Sensation Modulating Network of opponent dynamics.
If this is right
- Recursion and generativity reside in the modifiable dynamics of Negotiable Action Patterns.
- The self/world distinction is a structural feature of the wiring via the dual-signal property of self-modulatable action patterns.
- Grammar-grounded generativity arises as an architectural transition across the four-level hierarchy rather than an added layer.
- The cognitivism-4E impasse is resolved because recursion lives in modifiable dynamics while embodiment lives in the opponent substrate.
Where Pith is reading between the lines
- The seven testable registers listed in the appendix supply direct empirical checks on whether haltability suffices for intentional directedness.
- The body-wide broadcast network implies that attention can propagate across anatomical scales without requiring a separate centralized controller.
- The transition from Haltable to Negotiable patterns suggests a route by which public conventions could emerge directly from private bodily equilibria.
Load-bearing premise
The cognitive agent is the whole body organized at every anatomical scale by opponent dynamics built from sensation modulators paired into coordinated action zones routed by a body-wide broadcast network.
What would settle it
An observation or simulation of object-directed attention that requires a dedicated mechanism independent of co-activated equilibrium in bodily opponent dynamics.
Figures




















read the original abstract
Cognitive science remains split between cognitivism - which accounts for recursion and language but cannot ground formal symbols in meaning - and 4E approaches - which ground cognition in the body but rarely specify the body's architecture in enough detail to support generativity. We argue the impasse stems from an incomplete account of the embodied agent's architecture, and propose one: the Sensation Modulating Network (SMN), the cognitive agent conceived as the whole body, organized at every anatomical scale by opponent dynamics, built from Sensation Modulators that sense and act through one substrate, paired into Coordinated Action Zones routed by a body-wide broadcast network. Three commitments give the SMN its purchase. Haltability - the recruitment of antagonistic affordance into co-activated equilibrium - provides the architectural locus that object-directed phenomenology, in Husserl's sense, requires: opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness, with no module added on top. The dual-signal property of self-modulatable action patterns (SMAPs) makes the self/world distinction a structural feature of the wiring rather than a category the agent applies. And a four-level action-pattern hierarchy - Basal, Haltable, Negotiable, Transactional - gives a single trajectory from autonomic regularity to public conventionalization, locating the conditions for grammar-grounded generativity as architectural transitions. The SMN reconciles the cognitivism-4E debate: recursion lives in the modifiable dynamics of Negotiable Action Patterns, embodiment in the opponent substrate that supports them. A tentative formalism and eight predicted registers (seven testable, one hypothetical), with reference simulations, are given in an appendix.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes the Sensation Modulating Network (SMN) as the architecture of the embodied cognitive agent: the whole body organized at every scale by opponent dynamics built from Sensation Modulators paired into Coordinated Action Zones and routed by a body-wide broadcast network. It claims that haltability—the recruitment of antagonistic affordances into co-activated equilibrium—supplies the architectural locus for Husserlian object-directed phenomenology via the direct chain opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness, with no additional module required. Two further commitments are advanced: the dual-signal property of self-modulatable action patterns (SMAPs) makes the self/world distinction structural, and a four-level hierarchy (Basal, Haltable, Negotiable, Transactional) locates generativity in architectural transitions. A tentative formalism, eight predicted registers, and reference simulations appear in an appendix.
Significance. If the central architectural claim can be made rigorous, the work would supply a concrete, body-scale substrate that reconciles cognitivist recursion with 4E embodiment without postulating extra mechanisms, and would generate seven testable registers plus one hypothetical. The explicit credit given to the appendix formalism and simulations is a strength; the absence of free parameters in the core proposal is also noted.
major comments (3)
- [Haltability commitment paragraph] The section stating the three commitments (abstract and main text): the enablement chain 'opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness' is asserted as following directly from the SMN definitions, yet no derivation is supplied from the stated components (Sensation Modulators, Coordinated Action Zones, body-wide broadcast, or SMAP dual-signal property). This step is load-bearing for the claim that haltability alone grounds object-directed phenomenology with 'no module added on top.'
- [Haltability commitment paragraph] The definition of haltability itself (same section): haltability is introduced as 'the recruitment of antagonistic affordance into co-activated equilibrium' and then said to produce the phenomenology it is introduced to explain; the causal sequence therefore risks circularity rather than independent derivation from the opponent-dynamics wiring.
- [Appendix] Appendix (tentative formalism and reference simulations): if the formalism does not contain an explicit mapping from the four-level hierarchy and SMAP properties to the attention/directedness steps, the architectural-locus claim remains unclosed; the simulations are referenced but not shown to close this gap.
minor comments (1)
- [Four-level hierarchy paragraph] Notation for the four-level hierarchy (Basal, Haltable, Negotiable, Transactional) is introduced without a diagram or table summarizing transitions; a single table would improve readability.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review. The comments identify places where the central architectural claims require more explicit justification. We respond to each major comment below and indicate the revisions that will be made.
read point-by-point responses
-
Referee: [Haltability commitment paragraph] The section stating the three commitments (abstract and main text): the enablement chain 'opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness' is asserted as following directly from the SMN definitions, yet no derivation is supplied from the stated components (Sensation Modulators, Coordinated Action Zones, body-wide broadcast, or SMAP dual-signal property). This step is load-bearing for the claim that haltability alone grounds object-directed phenomenology with 'no module added on top.'
Authors: We agree that an explicit derivation of each link in the enablement chain from the SMN components was not supplied in the text. While the definitions of Sensation Modulators paired into Coordinated Action Zones and routed by the broadcast network imply that opponent dynamics permit co-activation of antagonistic affordances, that co-activation can reach equilibrium (halt), and that halt permits selective attention, this sequence was presented as following directly rather than derived step by step. We will add a dedicated subsection that derives each transition from the architectural elements, including the routing role of the body-wide broadcast network. revision: yes
-
Referee: [Haltability commitment paragraph] The definition of haltability itself (same section): haltability is introduced as 'the recruitment of antagonistic affordance into co-activated equilibrium' and then said to produce the phenomenology it is introduced to explain; the causal sequence therefore risks circularity rather than independent derivation from the opponent-dynamics wiring.
Authors: Haltability is defined independently as an architectural property arising from the opponent-dynamics wiring of Sensation Modulators in Coordinated Action Zones: the recruitment of antagonistic affordances into co-activated equilibrium. This definition does not presuppose phenomenology. The subsequent claim is that this property supplies the locus for object-directed phenomenology via the enablement chain. We will revise the relevant paragraphs to separate the architectural definition more clearly from the derived phenomenological consequences and to state explicitly that the definition itself is independent of the phenomenology it is later shown to enable. revision: partial
-
Referee: [Appendix] Appendix (tentative formalism and reference simulations): if the formalism does not contain an explicit mapping from the four-level hierarchy and SMAP properties to the attention/directedness steps, the architectural-locus claim remains unclosed; the simulations are referenced but not shown to close this gap.
Authors: The appendix formalism models the four-level hierarchy and SMAP dual-signal properties, and the reference simulations illustrate haltability, but we agree that an explicit mapping from these elements to the attention and directedness steps is absent and that the simulations do not demonstrate the full chain. We will expand the appendix to include this mapping, showing how the hierarchy and SMAP properties support the intermediate transitions, and will augment the simulation descriptions to address the relevant steps where the model permits. revision: yes
Circularity Check
Haltability is defined in terms of the equilibrium that is then said to ground the phenomenology
specific steps
-
self definitional
[Abstract]
"Haltability - the recruitment of antagonistic affordance into co-activated equilibrium - provides the architectural locus that object-directed phenomenology, in Husserl's sense, requires: opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness, with no module added on top."
Haltability is introduced by defining it as recruitment into co-activated equilibrium; the same concept is then asserted to enable the full chain to intentional directedness. The reduction is by construction: the explanatory mechanism (co-activated equilibrium producing attention and directedness) is already contained in the definition of the term that is said to derive the phenomenology.
full rationale
The paper's central claim is that haltability supplies the architectural locus for Husserlian object-directed phenomenology via the asserted chain (opponency enables co-activation, co-activation enables halt, halt enables attention, attention enables intentional directedness). The provided abstract defines haltability explicitly as 'the recruitment of antagonistic affordance into co-activated equilibrium' and immediately states that this supplies the required locus 'with no module added on top.' The enablement sequence is presented as following directly from the SMN architecture, yet no independent derivation from the definitions of Sensation Modulators, Coordinated Action Zones, or the broadcast network is exhibited; the explanatory power is embedded in the choice of definition. This is the self-definitional pattern. No self-citations, fitted predictions, or uniqueness theorems appear in the text. The appendix is referenced only for tentative formalism and simulations, without evidence that it supplies an external mapping. The result is therefore forced by the definitional construction rather than derived from independent benchmarks.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Cognitive science remains split between cognitivism and 4E approaches due to an incomplete account of embodied agent architecture.
- domain assumption The cognitive agent is the whole body organized at every anatomical scale by opponent dynamics.
invented entities (5)
-
Sensation Modulating Network (SMN)
no independent evidence
-
Haltability
no independent evidence
-
Sensation Modulators
no independent evidence
-
Coordinated Action Zones
no independent evidence
-
Self-modulatable action patterns (SMAPs)
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Adams, Stewart Shipp, and Karl J
Rick A. Adams, Stewart Shipp, and Karl J. Friston. Predictions not commands: Active inference in the motor system. Brain Structure and Function, 218: 0 611--643, 2013. doi:10.1007/s00429-012-0475-5
-
[2]
Micah Allen and Karl J. Friston. From cognitivism to autopoiesis: towards a computational framework for the embodied mind. Synthese, 195 0 (6): 0 2459--2482, 2018. doi:10.1007/s11229-016-1288-5
-
[3]
Bernard J. Baars. A Cognitive Theory of Consciousness. Cambridge University Press, Cambridge, 1988
1988
-
[4]
Bernard J. Baars. Global workspace theory of consciousness: Toward a cognitive neuroscience of human experience. Progress in Brain Research, 150: 0 45--53, 2005. doi:10.1016/S0079-6123(05)50004-9
-
[5]
Ruzena Bajcsy. Active perception. Proceedings of the IEEE, 76 0 (8): 0 966--1005, 1988. doi:10.1109/5.5968
-
[6]
Steps to an ecology of mind: Collected essays in anthropology, psychiatry, evolution, and epistemology
Gregory Bateson. Steps to an ecology of mind: Collected essays in anthropology, psychiatry, evolution, and epistemology. University of Chicago press, 2000
2000
-
[7]
The Brain's Sense of Movement
Alain Berthoz. The Brain's Sense of Movement. Harvard University Press, Cambridge, MA, 2000. Originally published in French as Le sens du mouvement, 1997
2000
-
[8]
Emilio Bizzi and Vincent C. K. Cheung. The neural origin of muscle synergies. Frontiers in Computational Neuroscience, 7: 0 51, 2013. doi:10.3389/fncom.2013.00051
-
[9]
Sarah-Jayne Blakemore, Daniel M. Wolpert, and Christopher D. Frith. Central cancellation of self-produced tickle sensation. Nature Neuroscience, 1 0 (7): 0 635--640, 1998. doi:10.1038/2870
-
[10]
Language Models are Few-Shot Learners
Tom B. Brown et al. Language models are few-shot learners. Advances in Neural Information Processing Systems, 33, 2020. arXiv:2005.14165
work page internal anchor Pith review Pith/arXiv arXiv 2020
-
[11]
Sean B. Carroll. Endless Forms Most Beautiful: The New Science of Evo Devo. W. W. Norton, 2005
2005
-
[12]
An outline of a theory of affordances
Anthony Chemero. An outline of a theory of affordances. Ecol. Psychol., 15: 0 181--195, 2003. ISSN 1040-7413. doi:10.1207/S15326969ECO1502_5
-
[13]
Syntactic structures
Noam Chomsky. Syntactic structures. Mouton, 1957
1957
-
[14]
Cortical mechanisms of action selection: the affordance competition hypothesis
Paul Cisek. Cortical mechanisms of action selection: the affordance competition hypothesis. Philosophical Transactions of the Royal Society B, 362: 0 1585--1599, 2007. doi:10.1098/rstb.2007.2054
-
[15]
The affordance competition hypothesis: The brain as a continuously predicting and action-selecting device
Paul Cisek. The affordance competition hypothesis: The brain as a continuously predicting and action-selecting device. Philosophical Transactions of the Royal Society B, 365 0 (1531): 0 1135--1145, 2010
2010
-
[16]
Paul Cisek and John F. Kalaska. Neural mechanisms for interacting with a world full of action choices. Annual Review of Neuroscience, 33: 0 269--298, 2010. doi:10.1146/annurev.neuro.051508.135409
-
[17]
Being There: Putting Brain, Body, and World Together Again
Andy Clark. Being There: Putting Brain, Body, and World Together Again. MIT Press, Cambridge, MA, 1997
1997
-
[18]
Supersizing the Mind: Embodiment, Action, and Cognitive Extension
Andy Clark. Supersizing the Mind: Embodiment, Action, and Cognitive Extension. Oxford University Press, 2008
2008
-
[19]
Trinity B. Crapse and Marc A. Sommer. Corollary discharge across the animal kingdom. Nature Reviews Neuroscience, 9: 0 587--600, 2008. doi:10.1038/nrn2457
-
[20]
Kathleen E. Cullen. Vestibular processing during natural self-motion: Implications for perception and action. Nature Reviews Neuroscience, 20: 0 346--363, 2019. doi:10.1038/s41583-019-0153-1
-
[21]
Kathleen E. Cullen, Jessica X. Brooks, and Soroush G. Sadeghi. How actions alter sensory processing: Reafference in the vestibular system. Annals of the New York Academy of Sciences, 1164: 0 29--36, 2009. doi:10.1111/j.1749-6632.2009.03866.x
-
[22]
Antonio R. Damasio. Descartes' Error: Emotion, Reason, and the Human Brain. Putnam, 1994
1994
-
[23]
Antonio R. Damasio. The Feeling of What Happens: Body and Emotion in the Making of Consciousness. Harcourt, 1999
1999
-
[24]
Antonio R. Damasio. Descartes' error revisited. J. Hist. Neurosci., 10: 0 192--194, August 2001. ISSN 0964-704X, 1744-5213. doi:10.1076/jhin.10.2.192.7250
-
[25]
Antonio R. Damasio. Self Comes to Mind: Constructing the Conscious Brain. Pantheon, 2010
2010
-
[26]
Antonio R. Damasio. Feeling & Knowing: Making Minds Conscious. Pantheon, 2021
2021
-
[27]
Consciousness and the Brain: Deciphering How the Brain Codes Our Thoughts
Stanislas Dehaene. Consciousness and the Brain: Deciphering How the Brain Codes Our Thoughts. Viking, 2014. ISBN 9780520274503
2014
-
[28]
Ezequiel A. Di Paolo. Autopoiesis, adaptivity, teleology, agency. Phenomenology and the Cognitive Sciences, 4 0 (4): 0 429--452, 2005. doi:10.1007/s11097-005-9002-y
-
[29]
Hubert L. Dreyfus. What Computers Can't Do: A Critique of Artificial Reason. Harper & Row, New York, 1972
1972
-
[30]
Hubert L. Dreyfus. What Computers Still Can't Do: A Critique of Artificial Reason. MIT Press, Cambridge, MA, 1992
1992
-
[31]
Dreyfus and Stuart E
Hubert L. Dreyfus and Stuart E. Dreyfus. Mind Over Machine: The Power of Human Intuition and Expertise in the Era of the Computer. Free Press, New York, 1986
1986
-
[32]
Anatol G. Feldman. Once more on the equilibrium-point hypothesis (lambda model) for motor control. Journal of Motor Behavior, 18: 0 17--54, 1986. doi:10.1080/00222895.1986.10735369
-
[33]
Anatol G. Feldman. Referent Control of Action and Perception: Challenging Conventional Theories in Behavioral Neuroscience. Springer, New York, 2015. doi:10.1007/978-1-4939-2736-4
-
[34]
Jerry A. Fodor. The Language of Thought. Harvard University Press, 1975
1975
-
[35]
Karl J. Friston. The free-energy principle: a unified brain theory? Nature Reviews Neuroscience, 11 0 (2): 0 127--138, 2010. doi:10.1038/nrn2787
-
[36]
Karl J. Friston. What is optimal about motor control? Neuron, 72 0 (3): 0 488--498, 2011. doi:10.1016/j.neuron.2011.10.018
-
[37]
Tom Froese and Ezequiel A. Di Paolo. The enactive approach: Theoretical sketches from cell to society. Pragmatics & Cognition, 19: 0 1--36, January 2011. ISSN 0929-0907, 1569-9943. doi:10.1075/pc.19.1.01fro
-
[38]
Di Paolo
Tom Froese and Ezequiel A. Di Paolo. Motivation, meaning, and the phenomenology of life. Phenomenology and the Cognitive Sciences, 10: 0 543--564, 2011
2011
-
[39]
How the Body Shapes the Mind
Shaun Gallagher. How the Body Shapes the Mind. Oxford University Press, Oxford, 2005
2005
-
[40]
Enactivist Interventions: Rethinking the Mind
Shaun Gallagher. Enactivist Interventions: Rethinking the Mind. Oxford University Press, 2017. Summary: Argues for enactive, embodied accounts of cognition, emphasizing how body and environment co-constitute experience; supports the shift from CNS-centric to body-as-network perspectives. Keywords: enactivism, embodiment, phenomenology, cognition
2017
-
[41]
Action and Interaction
Shaun Gallagher. Action and Interaction. Oxford University Press, 2021
2021
-
[42]
James J. Gibson. The Ecological Approach to Visual Perception. Houghton Mifflin, 1979
1979
-
[43]
Biological pattern generation: the lamprey as a model
Sten Grillner. Biological pattern generation: the lamprey as a model. Nature Reviews Neuroscience, 7 0 (1): 0 53--60, 2006
2006
-
[44]
Current principles of motor control, with special reference to vertebrate locomotion
Sten Grillner and Abdeljabbar El Manira. Current principles of motor control, with special reference to vertebrate locomotion. Physiological Reviews, 100 0 (1): 0 271--320, 2020. doi:10.1152/physrev.00015.2019. URL https://journals.physiology.org/doi/pdf/10.1152/physrev.00015.2019
-
[45]
David Ha and J\"urgen Schmidhuber. World models. Advances in Neural Information Processing Systems, 31, 2018. doi:10.5281/zenodo.1207631. arXiv:1803.10122
work page internal anchor Pith review Pith/arXiv arXiv doi:10.5281/zenodo.1207631 2018
-
[46]
Hermann Haken, J. A. Scott Kelso, and Herbert Bunz. A theoretical model of phase transitions in human hand movements. Biological Cybernetics, 51 0 (5): 0 347--356, 1985. doi:10.1007/BF00336922
-
[47]
Stevan Harnad. The symbol grounding problem. Physica D, 42 0 (1-3): 0 335--346, 1990. doi:10.1016/0167-2789(90)90087-6
-
[48]
Towards a theoretical foundation for morphological computation with compliant bodies
Helmut Hauser, Auke Jan Ijspeert, Rudolf Marcel F \"u chslin, Rolf Pfeifer, and Wolfgang Maass. Towards a theoretical foundation for morphological computation with compliant bodies. Biological Cybernetics, 105 0 (5--6): 0 355--370, 2011. doi:10.1007/s00422-012-0471-0
-
[49]
Hauser, Tecumseh W
Marc D. Hauser, Tecumseh W. Fitch, and Daniel J. Penn. The evolution of cognitive architecture: Reshaping the debate. Trends in Cognitive Sciences, 12 0 (3): 0 118--126, 2008
2008
-
[50]
Being and Time
Martin Heidegger. Being and Time. Harper & Row, New York, 1962. Translated by John Macquarrie and Edward Robinson; original Sein und Zeit published 1927
1962
-
[51]
The theory of event coding (tec): A framework for perception and action planning
Bernhard Hommel, Jochen M\"usseler, Gisa Aschersleben, and Wolfgang Prinz. The theory of event coding (tec): A framework for perception and action planning. Behavioral and Brain Sciences, 24 0 (5): 0 849--878, 2001. doi:10.1017/S0140525X01000103
-
[52]
Logical Investigations
Edmund Husserl. Logical Investigations. Routledge, London, 1900--1901. English translation by J. N. Findlay, Routledge 2001 (orig. Logische Untersuchungen, 1900--1901)
1900
-
[53]
Ideas Pertaining to a Pure Phenomenology and to a Phenomenological Philosophy
Edmund Husserl. Ideas Pertaining to a Pure Phenomenology and to a Phenomenological Philosophy. Kluwer, Dordrecht, 1913. English translation by F. Kersten, 1983 (orig. Ideen zu einer reinen Phänomenologie, 1913)
1913
-
[54]
Reafference and the origin of the self in early nervous system evolution
G \'a sp \'a r J \'e kely, Peter Godfrey-Smith, and Fred Keijzer. Reafference and the origin of the self in early nervous system evolution. Philosophical Transactions of the Royal Society B: Biological Sciences, 376 0 (1821): 0 20190764, 2021. doi:10.1098/rstb.2019.0764
-
[55]
Kandel, James H
Eric R. Kandel, James H. Schwartz, Thomas M. Jessell, Steven A. Siegelbaum, and A. J. Hudspeth. Principles of Neural Science. McGraw-Hill, New York, 5th edition, 2013. ISBN 978-0-07-139011-8
2013
-
[56]
J. A. Scott Kelso. Dynamic Patterns: The Self-Organization of Brain and Behavior. MIT Press, 1995
1995
-
[57]
Mark L. Latash. Evolution of motor control: From reflexes and motor programs to the equilibrium-point hypothesis. Journal of Human Kinetics, 19: 0 3--24, 2008. doi:10.2478/v10078-008-0001-2
-
[58]
A path towards autonomous machine intelligence
Yann LeCun. A path towards autonomous machine intelligence. Open Review preprint, 2022. Version 0.9.2, June 2022. https://openreview.net/pdf?id=BZ5a1r-kVsf
2022
-
[59]
7553, 436–444, https://doi.org/10.1038/nature14539
Yann LeCun, Yoshua Bengio, and Geoffrey Hinton. Deep learning. Nature, 521: 0 436--444, 2015. doi:10.1038/nature14539
-
[60]
Michael Levin. Morphogenetic fields in embryogenesis, regeneration, and cancer: Non-local control of complex patterning. BioSystems, 109 0 (3): 0 243--261, 2012. doi:10.1016/j.biosystems.2012.04.005
-
[61]
Michael Levin. Molecular bioelectricity: how endogenous voltage potentials control cell behavior and instruct pattern regulation in vivo. Molecular Biology of the Cell, 25 0 (24): 0 3835--3850, 2014. doi:10.1091/mbc.E13-12-0708
-
[62]
Michael Levin. Bioelectric networks: the cognitive glue enabling evolutionary and developmental scaling of goal-directedness in biology. Animal Cognition, 26: 0 1–20, 2023. doi:10.1007/s10071-023-01780-3. URL https://link.springer.com/article/10.1007/s10071-023-01780-3
-
[63]
Michael Levin and Giovanni Pezzulo. Endogenous bioelectric signaling networks: Exploiting voltage gradients for control of growth and form. Annual Review of Biomedical Engineering, 18: 0 243--268, 2016. doi:10.1146/annurev-bioeng-071114-040647
-
[64]
I of the vortex: From neurons to self
R R Llin \'a s. I of the vortex: From neurons to self . The MIT Press, Massachusetts, 2001. ISBN 9780262621632
2001
-
[65]
Variability, compensation, and modulation in neurons and circuits
Eve Marder. Variability, compensation, and modulation in neurons and circuits. Proceedings of the National Academy of Sciences, 108 0 (Suppl 3): 0 15542--15548, 2011. doi:10.1073/pnas.1010674108. URL https://www.pnas.org/doi/10.1073/pnas.1010674108
-
[66]
New insights from small rhythmic circuits
Eve Marder. New insights from small rhythmic circuits. Current Opinion in Neurobiology, 76: 0 102600, 2022. doi:10.1016/j.conb.2022.102600. URL https://www.sciencedirect.com/science/article/pii/S0959438822001040
-
[67]
Central pattern generators and the control of rhythmic movements
Eve Marder and Dieter Bucher. Central pattern generators and the control of rhythmic movements. Current Biology, 11 0 (23): 0 R986--R996, 2001. doi:10.1016/S0960-9822(01)00581-4. URL https://pubmed.ncbi.nlm.nih.gov/11728329/
-
[68]
Eve Marder and Ronald L. Calabrese. Principles of rhythmic motor pattern generation. Physiological Reviews, 76 0 (3): 0 687--717, 1996. doi:10.1152/physrev.1996.76.3.687. URL https://journals.physiology.org/doi/abs/10.1152/physrev.1996.76.3.687
-
[69]
Maturana and Francisco J
Humberto R. Maturana and Francisco J. Varela. Autopoiesis and Cognition: The Realization of the Living. Number 42 in Boston Studies in the Philosophy of Science. D. Reidel, 1980. ISBN 9027710155
1980
-
[70]
Homeobox genes and axial patterning
William McGinnis and Robb Krumlauf. Homeobox genes and axial patterning. Cell, 68 0 (2): 0 283--302, 1992. doi:10.1016/0092-8674(92)90471-N
-
[71]
Mehta, Diane Whitmer, Rodolfo Figueroa, Bryan A
Sachin B. Mehta, Diane Whitmer, Rodolfo Figueroa, Bryan A. Williams, and David Kleinfeld. Active spatial perception in the vibrissa scanning sensorimotor system. PLoS Biology, 5 0 (2): 0 e15, 2007. doi:10.1371/journal.pbio.0050015
-
[72]
Maurice Merleau-Ponty. Phenomenology of Perception. Routledge, London, England, April 2013. ISBN 9781135718602. doi:10.4324/9780203720714
-
[73]
R. Chris Miall and Daniel M. Wolpert. Forward models for physiological motor control. Trends in Neurosciences, 19 0 (10): 0 329--336, 1996. doi:10.1016/S0893-6080(96)00035-4
-
[74]
Cynthia F. Moss and Annemarie Surlykke. Probing the natural scene by echolocation in bats. Frontiers in Behavioral Neuroscience, 4: 0 33, 2010. doi:10.3389/fnbeh.2010.00033
-
[75]
Animal Evolution: Interrelationships of the Living Phyla
Claus Nielsen. Animal Evolution: Interrelationships of the Living Phyla. Oxford University Press, 3 edition, 2012. ISBN 9780199606023
2012
-
[76]
Action in perception
Alva Noë. Action in perception. MIT Press, 2004. ISBN 9780262140881
2004
-
[77]
J. Kevin O'Regan and Alva No \"e . A sensorimotor account of vision and visual consciousness. Behavioral and Brain Sciences, 24 0 (5): 0 939--1031, 2001. doi:10.1017/S0140525X01000115
-
[78]
James L. Peterson. Petri nets. ACM Computing Surveys (CSUR), 9 0 (3): 0 223--252, 1977. doi:10.1145/356698.356702
-
[79]
Navigating the affordance landscape: Feedback control as a process model of behavior and cognition
Giovanni Pezzulo and Paul Cisek. Navigating the affordance landscape: Feedback control as a process model of behavior and cognition. Trends in Cognitive Sciences, 20 0 (6): 0 414--424, 2016. doi:10.1016/j.tics.2016.03.013
-
[80]
How the Body Shapes the Way We Think: A New View of Intelligence
Rolf Pfeifer and Josh Bongard. How the Body Shapes the Way We Think: A New View of Intelligence. MIT Press, 2007
2007
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.